





Abstract
Cough is induced by inhaled prostacyclin analogues including treprostinil (TRE), and, at higher doses, treprostinil palmitil (TP), a prodrug of TRE. In this report, we have investigated mechanisms involved in TRE- and TP-induced cough, using a dry powder formulation of TP (TPIP) to supplement previous data obtained with an aqueous suspension formulation of TP (TPIS).
Experiments in guinea pigs and rats investigated the prostanoid receptor subtype producing cough and whether it involved activation of sensory nerves in the airways and vasculature. Experiments involved treatment with prostanoid, tachykinin and bradykinin receptor antagonists, a cyclooxygenase inhibitor and TRE administration to the isolated larynx or intravenously.
In guinea pigs, cough with inhaled TRE (1.23 µg·kg−1) was not observed with an equivalent dose of TPIP and required higher inhaled doses (12.8 and 35.8 µg·kg−1) to induce cough. TRE cough was blocked with IP and tachykinin NK1 receptor antagonists but not with EP1, EP2, EP3, DP1 or bradykinin B2 antagonists or a cyclooxygenase inhibitor. TRE administered to the isolated larynx or intravenously in rats produced no apnoea or swallowing, whereas citric acid, capsaicin and hypertonic saline had significant effects.
The mechanisms inducing cough with inhaled TRE likely involves the activation of prostanoid IP receptors on jugular C-fibres in the tracheobronchial airways. Cough induced by inhaled dry powder and nebulised formulations of TP occurs at higher inhaled doses than TRE, presumably due to the slow, sustained release of TRE from the prodrug resulting in lower concentrations of TRE at the airway sensory nerves.
Abstract
Cough induced by inhaled treprostinil and treprostinil palmitil involves the activation of prostacyclin (IP) receptors located on airway tachykinin nerves https://bit.ly/37sXz1I
Introduction
Treprostinil palmitil (TP) is a prodrug of the prostacyclin analogue treprostinil (TRE) that is designed to provide prolonged pulmonary vasodilation via an inhaled route of administration [1]. It is in development for the treatment of pulmonary arterial hypertension (PAH) and has been studied as an inhaled aqueous suspension (TPIS) and as an inhaled dry powder (TPIP) formulation. In preclinical studies, a single dose of TPIS provided long-acting inhibition of pulmonary vasoconstriction with detectable concentrations of TP remaining in the lungs for up to 24 h after administration [1, 2]. Consequently, the active drug TRE was also observed in plasma over the same extended time frame but at much lower levels compared to TPIS in the lung. In contrast, studies with inhaled TRE demonstrate its rapid elimination from the blood and lungs [1, 3, 4].
Cough and throat irritation are among the most common adverse events of inhaled TRE administered to PAH patients [3–6]. The mechanisms underlying this side-effect have not been previously elucidated. Conversely, TPIS does not cause cough at an equivalent inhaled dose to TRE that causes cough in both guinea pigs [1] and in human subjects [7, 8]. Here too, the factors involved in the lessened cough response to inhaled TP have not been defined.
In the present studies in guinea pigs and rats, we have investigated some of the potential mechanisms causing TRE-induced cough to identify the specific prostanoid receptor subtype(s) mediating TRE-induced cough, to elucidate the location and type of sensory nerve(s) producing this effect, to assess whether the presence of TRE in the blood was involved and to determine the actions of bradykinin or cyclooxygenase products that may be released by inhaled TRE. Furthermore, we evaluated whether inhaled TPIP acted like the other TP formulation (TPIS) and had a reduced propensity to cause cough compared to inhaled TRE, thereby distinguishing the contribution of the formulation from the prodrug itself. From these results, we have developed a hypothesis for why cough may be substantially lessened with administration of TP as either TPIS or TPIP. Some of these results have previously been reported in abstract form [7–11].
Materials and methods
Details of the methods and supporting data from preclinical studies in guinea pigs and rats can be found in the supplementary material. Also contained in the supplementary material is a description of the methods and results from a Phase 1 pharmacokinetic (PK) and safety evaluation for TPIS in healthy human volunteers.
Compounds and materials
TPIP was produced at Bend Research Inc. (Bend, OR, USA) as a formulated spray-dried powder and contained 1.5% of TP, 0.7% of DSPE-PEG2000, 68.2% of mannitol and 29.6% of l-leucine. A mannitol vehicle control was produced at Insmed Incorporated (Bridgewater Township, NJ, USA) and contained 0.7% of DSPE-PEG2000, 69.5% mannitol and 29.8% of l-Leucine. TRE and TP were obtained from Chirogate International (Taoyuan County, Taiwan, Republic of China). The following prostanoid, tachykinin and bradykinin antagonists were purchased: RO 1138452 (IP), ONO-8711 (EP1), PF-04418948 (EP2), L-798,106 (EP3), BW A868C (DP1) (Cayman Chemical, Ann Arbor, MI, USA), CP99994 (NK1) (Tocris Bioscience, Bristol, UK) and HOE 140 (bradykinin B2) (MedChem Express, Monmouth Junction, NJ, USA). Citric acid, capsaicin and meclofenamic acid were obtained from Sigma-Aldrich (St Louis, MO, USA). Phosphate-buffered saline (PBS) was acquired from Mediatech (Manassas, VA, USA). Details on preparation of solutions can be found in the supplementary material.
Animals
Male Hartley guinea pigs (300–460 g) and male Sprague Dawley rats (300–400 g) from Charles River Laboratories (St. Constant, Quebec, Canada) were housed in temperature-controlled (21°C) and humidity-controlled conditions with an acclimation period of 3–7 days. All experiments were performed in accordance with the Canadian Council for Animal Care (CCAC). Guinea pigs were acclimated to the plethysmograph once a day for a minimum of 3 consecutive days with a duration ranging from 30 min (for studies with TRE) up to 2 h (for TPIP).
Inhalation exposures in guinea pigs
Guinea pigs were placed inside a whole-body plethysmograph (Data Sciences International, Saint Paul, MN, USA) and exposed for 10 min to nebulised TRE or PBS using an Ultraneb Pro vibrating mesh nebuliser [1]. TPIP was administered for 15 min using a Vilnius Aerosol generator (VAG) (CH Technologies, Westwood, NJ, USA) [11]. A glass filter was attached to a plethysmograph outlet port, connected to a vacuum flow of 0.5 L·min−1 and the deposited drug (TRE or TP) collected over the duration of the aerosol exposure. The inhaled dose of TRE or TP was measured using an algorithm that has been previously described [12].
Cough in guinea pigs
Cough was measured from plethysmograph recordings, confirmed by manual observations, video recordings and from cough sounds using a microphone [1, 13]. Tidal volume, respiratory rate, minute ventilation and enhanced pause (Penh) were also measured. Baseline data were obtained over 15 min before drug exposure, followed by measurements every 5 min during the aerosol exposure and every 15 min after drug administration for up to 2 h post dose.
Prostanoid (IP, EP1, EP2, EP3 and DP1), tachykinin (NK1) and bradykinin (B2) antagonists and meclofenamic acid in guinea pigs
Guinea pigs were injected intraperitoneally with prostacyclin (IP), prostaglandin E2 (EP1, EP2 and EP3), prostaglandin D2 (DP1) and tachykinin NK1 receptor antagonists, meclofenamic acid or their respective vehicles 30 min before aerosolised TRE administration. The bradykinin B2 antagonist was administered for 10 min by inhalation with a pretreatment time of 10 min before aerosolised TRE administration. The doses and concentrations of these antagonists and their vehicles were based upon data from previous studies [14–17].
Laryngeal and cardiovascular reflexes in rats and guinea pigs
Rats and guinea pigs were anaesthetised with urethane (1.3 g·kg−1), and drugs were administered to the isolated laryngeal airway as previously described [18, 19]. Airflow through a lower tracheal catheter was measured with a pneumotachograph from which respiratory rate, tidal volume, minute ventilation, duration of expiration (TE) and apnoeic ratio (TE after nebulisation/TE before nebulisation) were measured [18–20]. A catheter was placed into the femoral artery for the measurement of the systemic arterial blood pressures (SAP). The number of swallows were enumerated by the presence of laryngeal elevation [20].
Nebulised test articles (PBS, citric acid and TRE) were delivered directly into the laryngeal catheter of rats for 20 s, and the apnoeic ratio, number of swallows and changes in SAP were measured. In guinea pigs, laryngeal administration of PBS or citric acid was delivered for 20 s and TRE was administered for 3 min.
For intravenous (i.v.) administration (TRE, capsaicin, PBS) to rats, the apnoeic ratio and the mean arterial blood pressure (MAP) were obtained before and after injection of the test articles as previously described [21].
Data analysis and statistics
In guinea pig cough studies, statistically significant differences between TRE, TPIP and their respective vehicles was determined with a one-way ANOVA followed by Tukey's post-test analysis using a GraphPadPrism 6 software package (GraphPad Software Inc., LaJolla, CA, USA). For studies involving administration of prostanoid, tachykinin and bradykinin antagonists and meclofenamic acid, significant differences between drug treatments or their respective vehicles were determined using an unpaired t-test. In rats, drug treatment effects on the apnoeic ratio, frequency of swallows and SAP were compared with the vehicle controls using a t-test with repeated measures. Statistical significance was denoted as p<0.05.
Results
TRE and TPIP on cough, ventilation and Penh in guinea pigs
Inhaled TRE produced cough at doses of 3.30 and 1.23 µg·kg−1, but not at a lower dose of 0.33 µg·kg−1 (figure 1a). Much higher doses of TPIP (12.8 and 35.8 µg·kg−1) were required to elicit cough compared to TRE (figure 1b), and at a dose of 2.3 µg·kg−1 no cough was observed. Cough was absent with the PBS and mannitol vehicles. The onset of cough with inhaled TRE occurred within the first 10 min of exposure, whereas the onset of cough with TPIP was delayed, occurring between 17 and 35 min after the start of the exposure for individual guinea pigs (figure 1c). Cough with both TRE and TPIP was identical to that described with prostaglandin E2 [22] and occurred in bouts of coughing with between 1 and 7 bouts per exposure. The cough response to TRE is illustrated in one guinea pig exposed to an inhaled TRE dose of 3.30 µg·kg−1 where 7 bouts of coughing were observed (figure 2).

Cough induced by increasing doses of treprostinil (TRE) and dry powder formulation of treprostinil palmitil (TPIP) in guinea pigs. Values are the mean±sem cough counts following exposure to a) TRE (n=4–10), b) TPIP (n=4–6) and c) the time to the first cough for both TRE and TPIP. *: p<0.05 compared to vehicle.

Plethysmograms of one guinea pig showing changes in inspiratory (Insp) and expiratory (Exp) airflows following exposure to treprostinil (TRE). a) Seven cough “bouts” evoked by a 10-min exposure to TRE (3.30 μg·kg−1). b) Expanded view of one of the cough bouts with 22 individual coughs enumerated.
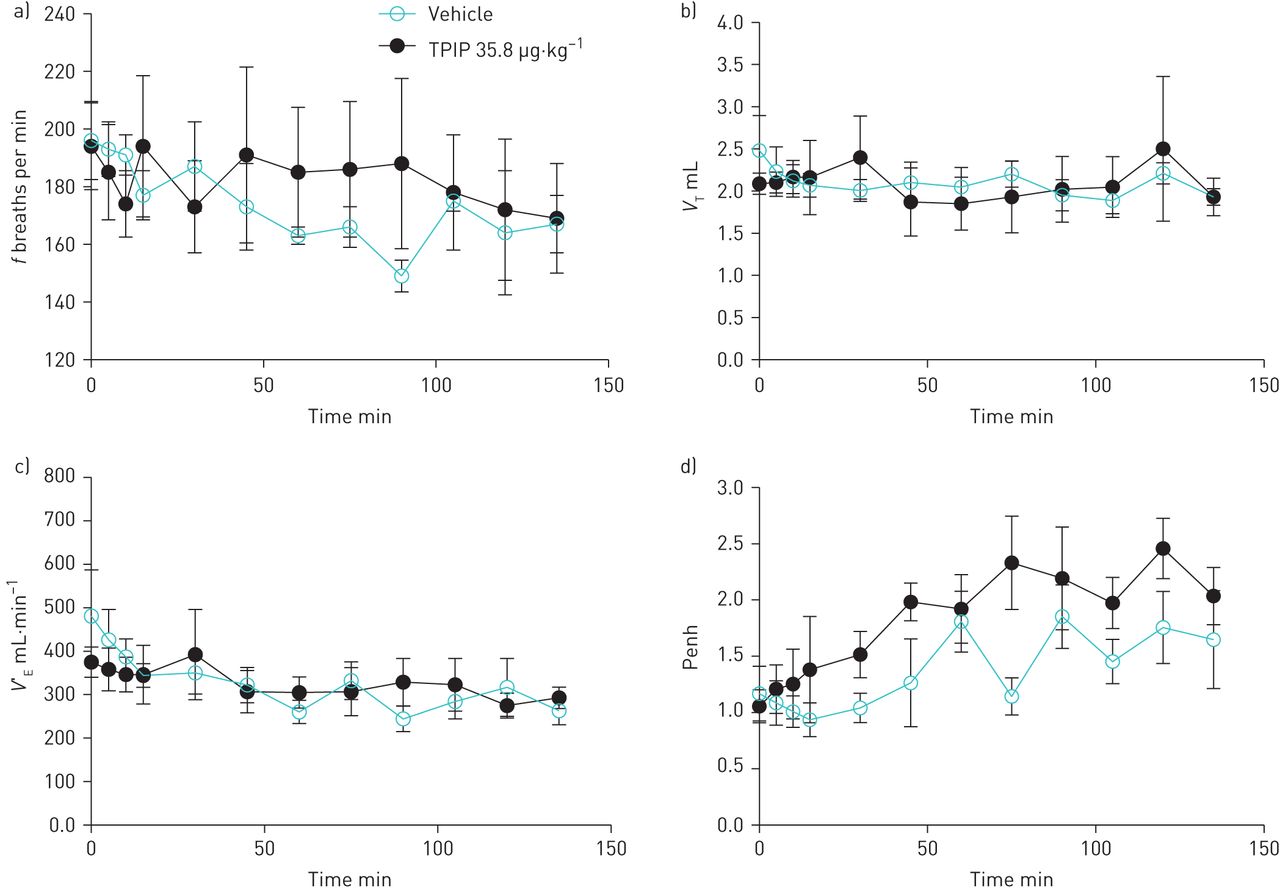
TPIP had no effects on ventilation and Penh at each inhaled dose. There was a slight but not statistically significant increase in Penh over time with TPIP at 35.8 µg·kg−1, but the same trend was observed with the mannitol vehicle (figure 3 and supplementary material). The magnitude of this increase in Penh with TPIP and the mannitol vehicle is much less than that observed with inhaled bronchoconstrictor agents such as citric acid and capsaicin that reach Penh values as high as 1000 along with laboured breathing (unpublished observations).

Effect of dry powder formulation of treprostinil palmitil (TPIP) and mannitol vehicle on ventilation and Penh in guinea pigs. Data are shown at baseline (time=0 min), during exposure to TPIP (time=0–15 min) and following TPIP exposure (time=15–135 min). TPIP was administered at an inhaled dose of 35.8 μg·kg−1. a) respiratory rate (f), b) tidal volume (VT), c) minute ventilation (V′E), and d) Penh (enhanced pause) over time. Values are the mean±sem (n=6 for TPIS and n=4 for mannitol vehicle).
Effect of prostanoid, tachykinin NK1 and bradykinin antagonists and meclofenamic acid on TRE cough in guinea pigs
TRE-induced cough was significantly (p<0.05) inhibited by the IP receptor antagonist RO 1138452 (10 mg·kg−1 intraperitoneally (i.p.)) but not with the EP1, EP2, EP3 and DP1 antagonists (figure 4a). In contrast, RO 1138452 (10 mg·kg−1 i.p.) had no effect on citric acid-induced cough, which induced 22±17 coughs in the presence of RO 1138452 and 28±17 coughs in the presence of the intraperitoneal vehicle control.

Effects of prostanoid receptor antagonists, a tachykinin NK1 receptor antagonist, a bradykinin B2 receptor antagonist and a cyclooxygenase inhibitor on treprostinil (TRE) cough in guinea pigs. The compound (blue bars) or their respective vehicles (white bars) were administered intraperitoneally (30 min) or by aerosol (10 min) prior to nebulised TRE (3.30 μg·kg−1). Graphs show the mean±sem effects against TRE-induced cough of a) the IP receptor antagonist (RO 1138452, 10 mg·kg−1 i.p., n=5), the EP1 receptor antagonist (ONO-8711, 10 mg·kg−1 i.p., n=5–6), the EP2 receptor antagonist (PF-04418948, 5 mg·kg−1 i.p., n=5), the EP3 receptor antagonist (L-798106, 10 mg·kg−1 i.p., n=5–7) and the DP1 receptor antagonist (BW A868C 10 mg·kg−1 i.p., n=5); and b) the NK1 receptor antagonist (CP99994, 10 mg·kg−1, i.p., n=5), the bradykinin B2 receptor antagonist (HOE 140, 1 mg·mL−1 delivered by aerosol, n=3) and the cyclooxygenase (COX) inhibitor (meclofenamic acid, 1 mg·kg−1 i.p., n=3). *: p<0.05 compared to vehicle.
TRE-induced cough was also significantly (p<0.05) inhibited by the tachykinin NK1 receptor antagonist CP99994 (10 mg·kg−1 i.p.), whereas treatment with the bradykinin B2 antagonist HOE 140 (1 mg·mL−1 delivered as an aerosol) or the cyclooxygenase inhibitor meclofenamic acid (1 mg·kg−1 i.p.) had no effects (figure 4b).
Laryngeal and cardiovascular reflexes with TRE in rats
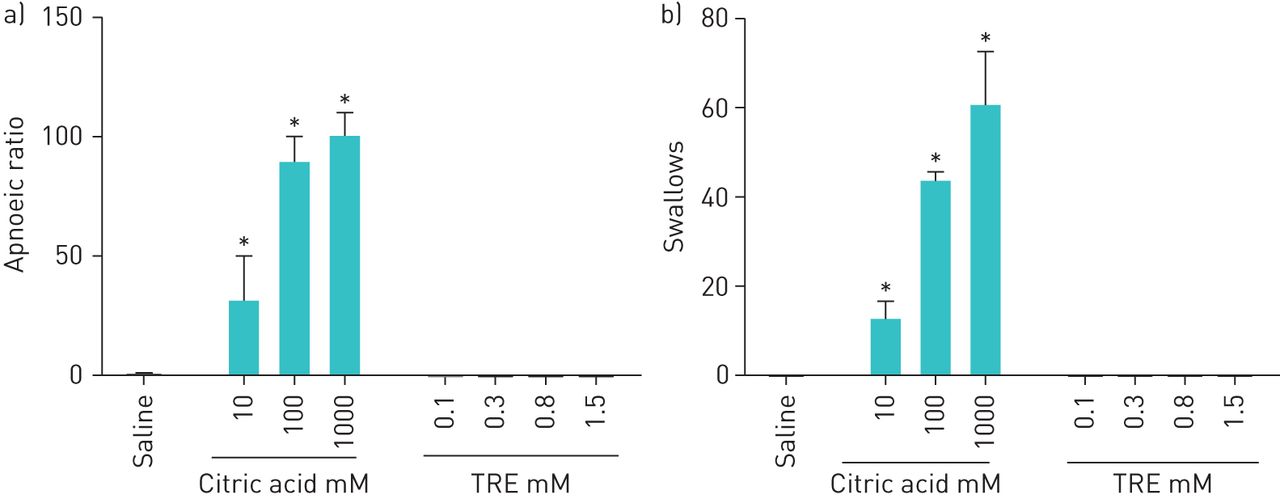
In anaesthetised rats, nebulised citric acid (10–1000 mM) administered directly into the larynx increased the apnoeic ratio and increased the frequency of swallows (figure 5a and b). Similar results were found with increasing concentrations of nebulised hypertonic saline (supplementary material). Nebulised TRE, at concentrations ranging from 0.1 to 1.5 mM, did not cause apnoea or increase the frequency of swallows (figure 5a and b). These concentrations are 2 to 30 times above the concentrations that trigger cough in conscious guinea pigs (figure 1).

Nebulisation of treprostinil (TRE) to the isolated laryngeal airway of rats produced no reflex effects, whereas nebulised citric acid produced concentration-dependent a) apnoea and b) increased the frequency of swallows. The apnoeic ratio was calculated as the TE treatment/TE baseline, where TE represents the duration of expiration. Values are the mean±sem (n=5 for saline, n=5 for citric acid, n=6 for TRE at 0.1 mM and n=2 for TRE at 0.3 to 1.5 mM). *: p<0.05 compared to saline.
Intravenous TRE (10–3000 ng·kg−1) had no effect on the apnoeic ratio and did not change the MAP (figure 6a and b), whereas a significant (p<0.05) increase in the apnoeic ratio and decrease in the MAP was seen with i.v. capsaicin (1250 ng·kg−1). The doses of i.v. TRE used for this evaluation exceed those that induce pulmonary vasodilation in rats [23].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Intravenous treprostinil (TRE) produced no reflex effects, whereas intravenous capsaicin produced a) apnoea and b) a decrease in mean arterial blood pressure (MAP). The apnoeic ratio was calculated as described in the legend to figure 5. Values represent the mean±sem (n=3). *: p<0.05 compared to saline.
Discussion
In clinical studies, cough and throat irritation are the most frequently reported adverse events with inhaled TRE and other inhaled prostanoid analogues such as iloprost [3–7]. To investigate the mechanisms involved with TRE cough, experiments were performed in guinea pigs, a species that responds to many of the tussive agents that cause cough in human subjects such as capsaicin, citric acid and prostaglandins [1, 13, 22, 24, 25]. Experiments were also performed in guinea pigs and rats to determine if TRE activates laryngeal or vascular reflexes that may be involved with cough following the local administration of TRE to the isolated larynx [18–20] or following intravenous administration [21]. The results from these studies demonstrate that TRE-induced cough involves the activation of IP receptors that are probably located on tachykinin-containing sensory nerves in the tracheobronchial airways. Furthermore, TPIP also induced cough in guinea pigs but required an approximately seven-fold (based upon the TRE equivalent inhaled dose) higher dose than inhaled TRE, consistent with observations for the TPIS formulation of TP.
Although TRE is classified as a prostacyclin analogue, it binds to several prostanoid receptors with a rank order of potency of: EP2>DP1>IP>EP1>EP3>EP4 and no binding (>10 µM) to the CRTH2 and FP receptors [1, 26]. To investigate the important prostanoid receptor subtype(s) involved with TRE-induced cough, guinea pigs were treated with potent and selective pharmacological antagonists for different receptor subtypes. TRE-induced cough was inhibited with RO 1138452, a selective IP antagonist [14], whereas the selective DP1, EP1, EP2 or EP3 antagonists [14–16] had no inhibitory effects. These results identify the IP receptor as the prostanoid receptor subtype mediating TRE-induced cough. Importantly, there were no effects with EP3 or DP1 antagonists, which are the prostanoid receptors involved with the cough response to prostaglandin E2 and prostaglandin D2 in guinea pigs [27, 28]. The selectivity of RO 1138452 for TRE-induced cough was confirmed in our study, as it had no effect against citric acid-induced cough.
Based upon previous studies that show prostacyclin analogues activate tachykinin-containing sensory nerves in various tissues, including airways [29–33], and from the data generated in the present study, it appears that cough induced by inhaled TRE likely involves the activation of IP receptors located on jugular C-fibres in the tracheobronchial airways. The fact that TRE-induced cough was blocked by CP99994, a selective tachykinin NK1 receptor antagonist [34], strongly suggests an involvement with the jugular C-fibres as neurokinins are expressed in jugular C-fibres but not nodose C-fibres [35]. The location of the sensory nerves is probably in the tracheobronchial airways, as topical administration of TRE to the isolated larynx of rats (figure 5) and guinea pigs ( supplementary material) produced no reflex effects such as apnoea, swallow and cough that occur with airway irritants such as citric acid or hypertonic saline [18–20]. Furthermore, intravenous TRE had no effects on the reflex apnoea and hypotension that occurs with intravenous irritants in rats [21], which eliminates a vascular site of action. There were no changes in respiratory rate or Penh and no evidence of bronchoconstriction with inhaled TPIP or TRE in guinea pigs (figure 3 and supplementary material), which likely rules out the involvement of mechanosensitive airway nerves or nodose C-fibres [35, 36]. Furthermore, cough induced by inhaled TRE did not involve the secondary release of mediators such as bradykinin or products from cyclooxygenase metabolism, as treatment with the bradykinin antagonist (HOE 140) or a cyclooxygenase inhibitor (meclofenamic acid) had no effects. In summary, these results strongly suggest that TRE-induced cough involved the activation of IP receptors on jugular C-fibres in the tracheobronchial airways, a conclusion that is consistent with previously published reports on the activation of non-adrenergic, non-cholinergic neurotransmission by non-prostanoid prostacyclin mimetics via activation of the IP1-receptor subtype [33, 37].
As previously mentioned, there is excellent translation between humans and guinea pigs for cough induced by a variety of different respiratory irritants including capsaicin, citric acid and prostaglandin E2 [1, 13, 22, 24, 25]. The results from this study indicate that inhaled TRE can be added to the list of tussive agents that show excellent translation between guinea pigs and humans for cough, as TRE produces cough in both guinea pigs [1, 9, 10] and humans [3, 4, 7], and experiments with the nebulised TP prodrug (TPIS) in both guinea pigs [1] and humans (supplementary material) demonstrate that TP can cause cough, but only at significantly higher doses relative to TRE. Though inhaled mannitol, an excipient of TPIP, is a known tussive agent in humans [38], we ruled out a contribution of a mannitol effect on the response to TPIP as the mannitol vehicle produced no cough. The fact that both TPIS and TPIP produced cough at higher inhaled doses compared to TRE suggests it is unlikely that there were protective effects from these different formulations. Rather, the difference is most likely due to the relatively slow conversion rate of the prodrug to TRE resulting in the markedly lower peak concentrations of TRE for the same molar dose of TP. It remains to be seen if TPIP produces cough at a higher inhaled dose than inhaled TRE in humans and whether cough occurs in delayed bouts of coughing as was seen in guinea pigs.
There are several limitations to the conclusions reached in this study. First, the mechanistic studies were performed in guinea pigs, so extrapolation of the results to humans is speculative. However, as discussed, there is excellent translation between guinea pigs and humans for the induction of cough by respiratory irritants, including TRE [1, 13, 22, 24, 25]. Furthermore, tachykinin receptor antagonists are now commercially available [39, 40], so it should be a straightforward task to determine if a tachykinin NK1 receptor antagonist can block TRE cough in humans. Another potential limitation in this study pertains to studies in human subjects in which the overall plasma TRE exposure (area under the concentration–time curve) with TPIS (85 µg) was less than TRE (54 µg), even though the inhaled TRE doses were identical on a molar basis (after conversion of TP to TRE for TPIS). Therefore, the bioequivalence of TPIS and TRE could be in question. However, in guinea pigs and dogs, a lower incidence of cough occurs with TPIS compared to inhaled TRE [1], so we believe the lower level of respiratory adverse events with TPIS in humans (supplementary material) likely reflects the unique pharmacokinetics of the prodrug, which delays presentation of TRE to the sensory nerves in the lungs and results in a significantly lower level of peak TRE exposure. Preclinical data with TPIP have found similar results of long-acting pulmonary vasodilation and a similarly low maximum plasma TRE concentration with sustained concentrations of TP in the lungs over 12 to 24 h (unpublished observations), so future studies of TPIP in humans should prove very enlightening should they confirm the findings of the preclinical data with respect to the cough response to equipotent doses of TP and TRE.
In conclusion, the results from this study suggest that activation of prostanoid IP receptors present on tachykinin-containing jugular C-fibres in the tracheobronchial airways mediate TRE-induced cough. As it appears that there is excellent translation between guinea pigs and humans in the cough response to inhaled TRE, it is likely that TPIP will behave in the same manner as TPIS and be associated with less cough in humans than inhaled TRE.
Supplementary material
Supplementary Material
Please note: supplementary material is not edited by the Editorial Office, and is uploaded as it has been supplied by the author.
Supplementary material 00592-2020.SUPPLEMENT
Footnotes
This article has supplementary material available from openres.ersjournals.com.
Conflict of interest: R.W. Chapman is an employee of Insmed Incorporated.
Conflict of interest: M.R. Corboz is an employee of Insmed Incorporated.
Conflict of interest: C. Fernandez is an employee of Insmed Incorporated.
Conflict of interest: E. Sullivan is an employee of Insmed Inc.
Conflict of interest: A. Stautberg is an employee of Insmed Incorporated.
Conflict of interest: A.J. Plaunt is an employee of Insmed Incorporated. In addition, he has patents pending or issued (patent numbers US 10,010,518, US 10,526,274, US 9,255,064 and US 9,469,600) related to treprostinil palmitil and methods of treatment via administration of treprostinil palmitil.
Conflict of interest: D.M. Konicek is an employee of Insmed Incorporated. In addition, she has patents pending or issued (patent numbers US 10,010,518, US 10,526,274, US 9,255,064 and US 9,469,600) related to treprostinil palmitil and methods of treatment via administration of treprostinil palmitil.
Conflict of interest: V. Malinin is an employee of Insmed Incorporated. In addition, he has patents pending or issued(patent numbers US 10,010,518, US 10,526,274, US 9,255,064 and US 9,469,600) related to treprostinil palmitil and methods of treatment via administration of treprostinil palmitil.
Conflict of interest: Z. Li is an employee of Insmed Incorporated. In addition, he has patents pending or issued(patent numbers US 10,010,518, US 10,526,274, US 9,255,064 and US 9,469,600) related to treprostinil palmitil and methods of treatment via administration of treprostinil palmitil.
Conflict of interest: D. Cipolla is an employee of Insmed Incorporated.
Conflict of interest: W. Perkins is an employee of Insmed Incorporated. In addition, he has patents pending or issued(patent numbers US 10,010,518, US 10,526,274, US 9,255,064 and US 9,469,600) related to treprostinil palmitil and methods of treatment via administration of treprostinil palmitil.
- Received August 17, 2020.
- Accepted November 23, 2020.
- Copyright ©ERS 2021
This article is open access and distributed under the terms of the Creative Commons Attribution Non-Commercial Licence 4.0.
References










