





Abstract
Lung cancer is one of the main causes of death worldwide. Published data show the use of interferons (IFNs) in treating lung tumours. IFNs also have potential for their antiproliferative, antiangiogenic, immunoregulatory and proapoptotic effects. IFN-γ functions as an anticancer agent against various forms of cancer. This study aimed to investigate the effect of IFN-γ liposome (nano) on peripheral lymphocytes from 20 individuals in each group: lung cancer patients compared to healthy individuals. The effectiveness of IFN-γ liposome against oxidative stress was also evaluated in this study. A concentration of 100 U·mL−1 of IFN-γ liposome was used to treat the lymphocytes in the Comet and micronucleus assays based on the preliminary test for the optimal dose. The lymphocytes from lung cancer patients presented with higher DNA damage levels than those of healthy individuals. In healthy individuals, IFN-γ liposome did not cause any DNA damage in the lymphocytes. Also, it caused a significant reduction in DNA damage in the lymphocytes from lung cancer patients in both the Comet and micronucleus assays. The 100 U·mL−1 of IFN-γ liposome significantly reduced the oxidative stress caused by H2O2 and appeared to be effective in both groups using the Comet and micronucleus assays. Results from both Comet and micronucleus assays were consistent. The data obtained indicated that IFN-γ in both forms (IFN-γ bulk and IFN-γ nanoliposome) may potentially be effective for the treatment of lung cancer and showed the ability of IFN-γ liposome to reduce DNA damage more than the bulk form.
Abstract
A research article on the effect of bulk and nanoliposome interferon-γ on lymphocytes from patients with lung cancer compared to healthy controls: assessing the ability of IFN-γ liposome to reduce DNA damage more than the bulk form https://bit.ly/3wvfSyY
Introduction
The term lung cancer encompasses epithelial cancers in the mucosa of the bronchi and sometimes in the parenchyma of the lung, which exists in the trachea (windpipe), bronchi (airways) or lung air sacs (alveoli) [1, 2]. Lung cancer accounts for roughly 13% of annual cancer cases worldwide. It is also the second most common type of cancer in both males and females within the UK, although its incidence is higher in males than females [3]. In the UK, lung cancer is the leading reason for cancer death, with approximately 35 000 deaths annually. The diagnostic peaks for lung cancer are between the ages of 73 and 84 years [4], and the overall 5-year survival rate is less than 10%, mainly because the majority of patients present with a late phase of the disease, a point at which treatment has little impact on survival [5]. This cancer could be developed or extended to the airways, resulting in symptoms such as cough, airway obstruction and haemoptysis. The tumour can metastasise to the thorax area, causing compression. It might also invade the chest wall and extend further, via the hilar, mediastinal and supraclavicular nodes [6]. Furthermore, it enhances transformations in the central or peripheral nervous system, leading to anorexia and causing a disturbance in hormone production. Cancer metastasises via the blood to different parts of the body, specifically to the liver, brain, adrenal gland and axial skeleton [7].
Studies have shown that cigarette smoking is the main risk factor in lung cancer patients. Current smokers have a 15-fold increase in risk of death from lung cancer compared with lifelong nonsmokers [8]. On the other hand, this risk decreases significantly in individuals who cease smoking before middle age [9]. The risk of lung cancer is also elevated following exposure to environmental tobacco smoke (ETS). An estimated 14–15% of lung cancers are caused by exposure to ETS [10]. 85% of lung cancer patients have nonsmall cell lung cancer (NSCLC), including adenocarcinoma, adenosquamous cell carcinoma, squamous cell carcinoma and large cell carcinoma.
IFN-γ is produced mainly by Th1 CD4+ T cells, CD8+ T cells and natural killer (NK) cells. Additionally, recent studies have indicated that other cells such as B cells, natural killer T (NKT) cells and professional antigen-presenting cells (APCs) may also be a source of IFN-γ. This secretion is controlled primarily by IL-12 and IL-18 [11]. IFN-γ has major antiviral, immunoregulatory and antitumour roles, through interaction with specific cell-surface receptors, including four transmembrane polypeptide receptor complexes: two ligand-binding chains (IFN-γR1) and two signal-transducing chains (IFN-γR2), which are required for signal transduction [12]. The most critical transduction pathway stimulated by IFN-γ is the Janus kinase (Jak)/signal transducer and activator of transcription 1 (STAT1) pathway. This contributes to the antiproliferative effects of IFN-γ on different cell types, as it has been elucidated that both pathways have the potential to inhibit tumour progression and kill pathogen-infected cells [13]. This is done by binding two IFN-γ R2 subunits with two IFN-γ-bound IFN-γ R1 chains [14]. After the receptor complex association is induced, Jak1 and Jak2 activate and phosphorylate Tyr440 of IFN-γ R1, forming a docking site for STAT1. STAT1 is activated by phosphorylation at Tyr701, leading to homodimerisation of STAT1 and nuclear translocation. The STAT1 dimers interact with specific DNA sequences called IFN-γ-activated sequences (GAS), present in the promoter regions of IFN-stimulated genes, and responsible for regulating their transcription [15, 16]. The entirety of these mechanisms are presented in figure 1 [17].
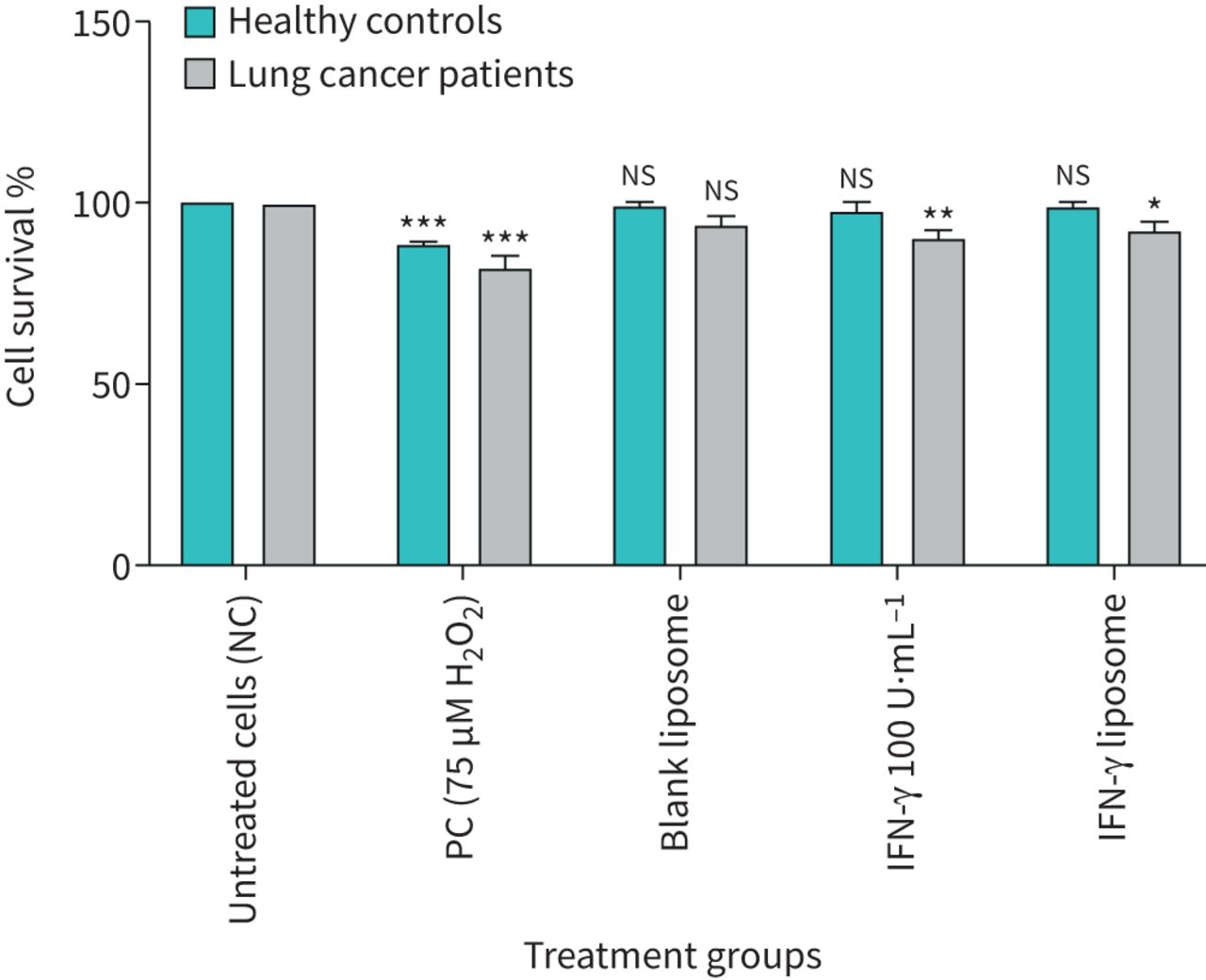
Per cent survival in 10 × 104 cells of various test treatments in lymphocytes from healthy individuals and lung cancer patients using CCK-8 assay. Error bars show mean values±se, n=3. NC: nucleated cell; PC: positive control; IFN: interferon; ns: nonsignificant. *: p<0.05; **: p<0.01; ***: p<0.001.
The transcriptional activity of STAT1 is stimulated through kinases. These kinases are mitogen-activated protein kinase (MAPK), protein kinase C (PKC) and phosphoinositide 3-kinase (PI3K)/AKT, which phosphorylates STAT1 in the transactivation domain [17]. The mechanism of the STAT1 pathway plays a positive role by regulating the expression of genes encoding several antiproliferative and proapoptotic molecules, such as IFN regulatory factor 1 (IRF-1). However, if IRF-1 is inhibited, the IFN-γ activates the proliferative signals. On the other hand, when the IRF-1 is elevated, the IFN-γ stimulates apoptotic signals [18, 19]. In mice, those with deficient IFN-γ spontaneously develop lung epithelial malignancies and lymphoma. Furthermore, mice with a deficient IFN-γ receptor and STAT1 manifest tumour growth following chemical carcinogen treatment, which emphasises the ability of IFN-γ to act as an anticancer agent [20]. Furthermore, preliminary research indicates that IFN-γ, given as a single agent, has a measurable effect as an antitumour agent in advanced NSCLC [21].
Liposomes are artificial vesicles made from single or multiple phospholipid layers (lamellas) with spherical, self-assembled shapes. Their sizes typically range from tens of nanometres up to hundreds of micrometres, depending on their preparation methods [22, 23]. They can also be made from nontoxic surfactants, cholesterols and membrane proteins [24]. Liposomes have been used extensively as drug delivery carriers and utilised for biomedical and biotechnological purposes [25]. The encapsulation of the drug into liposomes can reduce the toxicity, enhance the pharmacokinetics of the drug and result in more efficient therapeutic effects [26].
Moreover, liposomes can increase bioavailability and improve the solubility of poorly water-soluble drugs [5]. A recent study adopted the IFN-γ liposome-based drug delivery method by modifying it with cyclic peptides in order to initiate an antifibrotic effect for the therapy of hepatic fibrosis in the rat. It was found that the sterically modified IFN-γ liposomes have a greater antifibrotic effect of activating the hepatic stellate cells by 7.24-fold and 2.95-fold compared to free IFN-γ, with fewer adverse side-effects [27].
The primary objective of this research was to study the DNA protective effect of IFN-γ bulk and IFN-γ liposome on lymphocytes from lung cancer patients compared to healthy individuals.
In this study, the effect of IFN-γ bulk and IFN-γ liposome on DNA damage was detected in peripheral blood lymphocytes of lung cancer patients and healthy individuals by using the Comet micronucleus assay. The Comet assay was used in this study due to its sensitivity and simplicity for analysis of the genotoxicity of DNA in lymphocytes as presented in previous studies [28–30].
Materials and methods
Materials
IFN-γ (≥98% purity; Cat No. 17001), Mitomycin C (MMC) (Cat No. M0503) and fetal bovine serum (FBS) (Cat No. F7524) were purchased from Sigma-Aldrich (Gillingham, UK). RPMI 1640 Medium (RPMI-1640) (Cat No. R8758), phytohaemagglutinin (PHA) (Cat No. L1668) and penicillin–streptomycin solution (Cat No. P4333) were also obtained from Sigma-Aldrich. All other chemicals used in the different tests were from Sigma-Aldrich Company Ltd. (Sigma Chemical Ltd., Gillingham, UK). Prior to using the IFN-γ, the lyophilised powder was reconstituted in double distilled water to form the stock solution. It was then diluted in RPMI-1640 Medium with 10% FBS and stored at −20°C.
The dose–response experiments were carried out to determine the optimal doses of IFN-γ bulk and liposome and H2O2 used throughout the study. A fixed dose of both forms (100 U·mL−1 of IFN-γ bulk and 100 U·mL−1 of IFN-γ liposome) were used as treatment. The classic genotoxic compound hydrogen peroxide (H2O2) (75 μM) was used as a positive control (PC).
Preparation and characterisation of liposomes
Liposomes were prepared using the thin film rehydration method. 1,2-Dipalmitoyl-sn-glycero-3-phospho-rac-(1-glycerol) sodium salt (DPPG), 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine (DOPE) and cholesterol were dissolved in dichloromethane (DCM) and methanol (3:1 v/v). The solution was then transferred to a rotary evaporator with a 25 mL round-bottomed flask. The organic solvent mixture was evaporated at 40°C under vacuum for 6 h, to ensure no traces of the organic solvents were left. The thin film was then hydrated by 10 mL of PBS solution of IFN-γ for 1 h at 60°C using a bath sonicator (150 W). The sample was then subjected to four repeated freeze–thaw cycles (freezing at −20°C and thawing at 60°C in the bath sonicator). Furthermore, the blank liposome was prepared from the same components without adding the IFN-γ.
Average size and polydispersity index (PDI) of the liposome preparations were determined by dynamic light scattering (DLS) using Zetasizer ZS (Malvern Instruments, UK). All measurements were performed in triplicate.
Ethical approval
Ethical approval was obtained to execute the Comet and micronucleus assays for the study of IFN-γ and IFN-γ liposome. This study received ethical approval from Leeds East Research Ethics Committee (REC number: 12/YH/0464), the University of Bradford Research Ethics Sub-Committee on Research in Human Subjects (Ref: 0405/8) and the Research Support and Governance Office, Bradford Teaching Hospitals, NHS Foundation (Ref: RE DA 1202).
Sample preparation and Comet assay
After obtaining consent from healthy nonsmoking volunteers and lung cancer patients, whole blood samples were collected by venepuncture in 9 mL lithium heparin-coated tubes and clearly labelled for ease of identification. Samples were diluted in a ratio of 1:1 with RPMI-1640 Medium, and then mixed with 10% dimethyl sulfoxide (DMSO). The diluted blood solution was divided and transferred to labelled 1.5 mL Eppendorf® tubes, which were closed tightly and stored at −80°C.
The Comet assay was prepared according to [31, 32]. In brief, 100 μL of whole blood samples were incubated for 30 min at 37°C with different test agents, to make the final volume of 1000 μL with RPMI-1640 Medium (with sodium bicarbonate, without L-glutamine, liquid, sterile-filtered, suitable for cell culture). The test agents included a PC of 75 μM H2O2, blank liposome IFN-γ 100 U·mL−1 and IFN-γ liposome in order to test the effect of IFN-γ in a highly oxidising environment; 100 U·mL−1 of IFN-γ and IFN-γ liposome were treated in the presence of 75 mM H2O2. An untreated sample was used as a negative control. The samples were then incubated at 37°C for 30 min. Following incubation, the samples were centrifuged at 3000 rpm for 5 min. The method used thereafter was followed as described in [28–30].
The slides were coded before scoring; per slide, 100 nuclei were scored using a fluorescent microscope at 20× magnification. This was connected to a CCD camera using Komet 6 software and Kinetic Imaging (Andor Technology Ltd, Belfast). Olive tail moment (OTM) and % tail DNA were used simultaneously to decrease variability in the results [33].
The cytokinesis block micronucleus (CBMN) assay
Whole blood (500 μL) was added to a T25 cm2 Corning culture flask containing 4.5 mL RPMI-1640 Medium supplemented with 1% penicillin–streptomycin, 15% FBS and 25 mM HEPES and L-glutamine with end concentrations of 15% and 1%, respectively, followed by 100 μL of phythaemagglutinin (PHA) (2.5%). In the next 24 h, 50 μL of blank liposome was added to the negative control. Two positive controls were used with 10 μL of MMC (0.4 μM) used as PC 1, and 50 μL of H2O2 served as PC 2. 50 μL of 100 U·mL−1 IFN-γ and IFN-γ liposome were added to their respective flasks and the final flask contained IFN-γ liposome and H2O2. 50 μL of cytochalasin-B (6 μg·mL−1, Sigma) was added to each flask and cultured at 37°C in the presence of 5% CO2 for 44 h. The CBMN procedures were performed as described by Fenech and coworkers [34, 35].
Numerous cytological scoring parameters were used to assess DNA damage. Micronuclei (MNi) were scored from both binucleated cells (BiNCs) and mononucleated cells (MonoNCs). Nucleoplasmic bridges (NPBs) and nuclear buds (NBUDs) were only scored for BiNC [36]. The nuclear division index (NDI) was calculated using the equation NDI=M1+2(M2)+3(M3)/N, where M1=mononucleated cells, M2=binucleated cells, M3=multinucleated cells and N=total number of viable cells scored [35].
Cell viability
The viability of cells was detected after 30 min of incubation of cells with different concentrations of treatments. Lymphocytes were isolated from whole blood using ficoll-paque as described in [37]. Cell viability was then determined by the trypan blue staining method described in [38] and the Cell Counting Kit-8 (CCK-8) method [39]. The concentrations with cell viability of ≥75% were used in all experiments [40].
Statistical analysis
The final data were expressed as mean values with standard errors, the results were analysed using one-way ANOVA with Dunnett's multiple comparisons test using GraphPad Prism 8.1.2. p-values were considered significant at p<0.05, p<0.01, p<0.001.
Results
The viability of lymphocytes from healthy individuals and lung cancer patients was assessed after treatment with chemicals used in this study was between 75% and 85% using trypan blue dye exclusion. The viability of lymphocytes from healthy individuals and lung cancer patients treated with different treatment groups was also confirmed using the CCK-8 assay ≥75 after 24 h treatment (figure 1).
Comet assay: IFN-γ liposome versus bulk
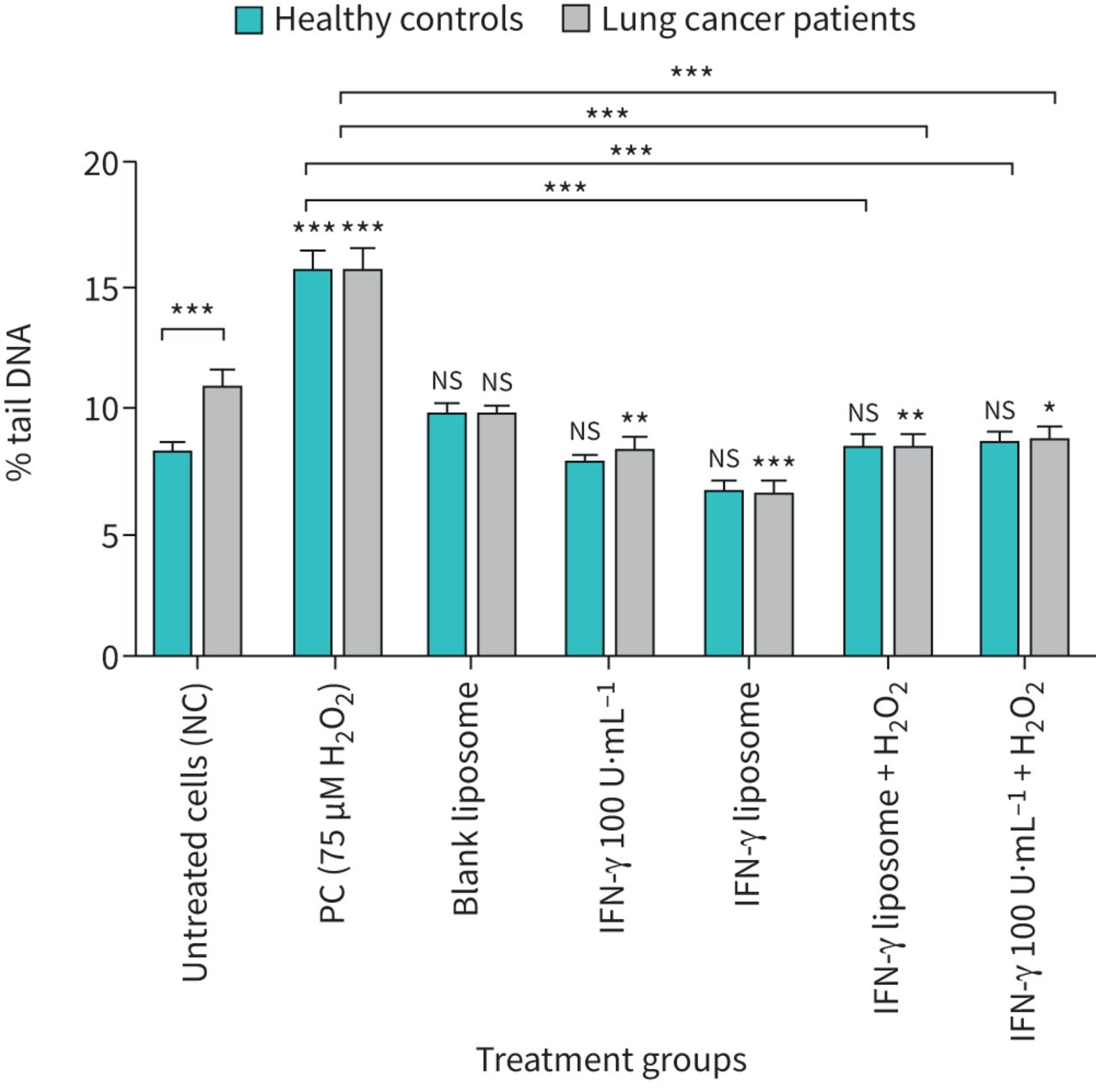
The data from the Comet assay results showed that lung cancer patient lymphocytes were more susceptible to DNA damage than healthy individuals, in % tail DNA and OTM, when compared with groups that received no treatment (p<0.001) as presented in table 1, figures 2 and 3. A significant increase in % tail DNA and OTM in lymphocytes from healthy volunteers and lung cancer patients after exposure to PC (75 μM H2O2) (p≤0.001) (figures 2 and 3).
The mean values of experiments on blood samples from 20 healthy individuals and 20 lung cancer patients expressed as OTM and % tail DNA

DNA damage measured as mean olive tail moment before and after treatment with bulk and liposome forms of interferon (IFN)-γ in human lymphocytes from healthy individuals and lung cancer patients in the Comet assay. n=20 in each group. NC: nucleated cells; PC: positive control; ns: nonsignificant. *: p<0.05; **: p<0.01; ***: p<0.001.

DNA damage measured as mean % tail DNA before and after treatment with bulk and liposome forms of interferon (IFN)-γ in human lymphocytes from healthy individuals and lung cancer patients in the Comet assay. n=20 in each group. NC: nucleated cells; PC: positive control; ns: nonsignificant. *: p<0.05; **: p<0.01; ***: p<0.001.
Lymphocytes obtained from 20 healthy individuals (figures 2 and 3) showed that 100 U·mL−1 IFN-γ bulk and IFN-γ liposome was clearly unaffected in lymphocyte cells when measuring % tail DNA and OTM compared to untreated cells. In contrast, the H2O2-induced DNA damage was significantly reduced by 100 U·mL−1 IFN-γ bulk and IFN-γ liposome in lymphocytes from healthy individuals compared to the PC (75 μM H2O2) (p≤0.001). In contrast, IFN-γ liposome was more effective in reducing the DNA damage than IFN-γ bulk, as presented in table 1.
Lymphocytes from lung cancer patients (n=20) also showed a significant decrease in % tail DNA and OTM from 10.97% (% tail DNA) and 2.18 (OTM) compared to the untreated control groups to 8.42% (% tail DNA) and 1.38 (OTM), respectively, when cells were treated with IFN-γ bulk whereas the reduction in % tail DNA and OTM was more effective by using IFN-γ liposome, which showed a significant decrease in % tail DNA 6.65% and 1.07 (OTM). Furthermore, cells treated with IFN-γ bulk and IFN-γ liposome co-supplemented with 75 μM H2O2 showed significant decreases in OTM and % tail DNA compared to the PC (75 μM H2O2) (p≤0.001).
CBMN: effect of IFN-γ liposome
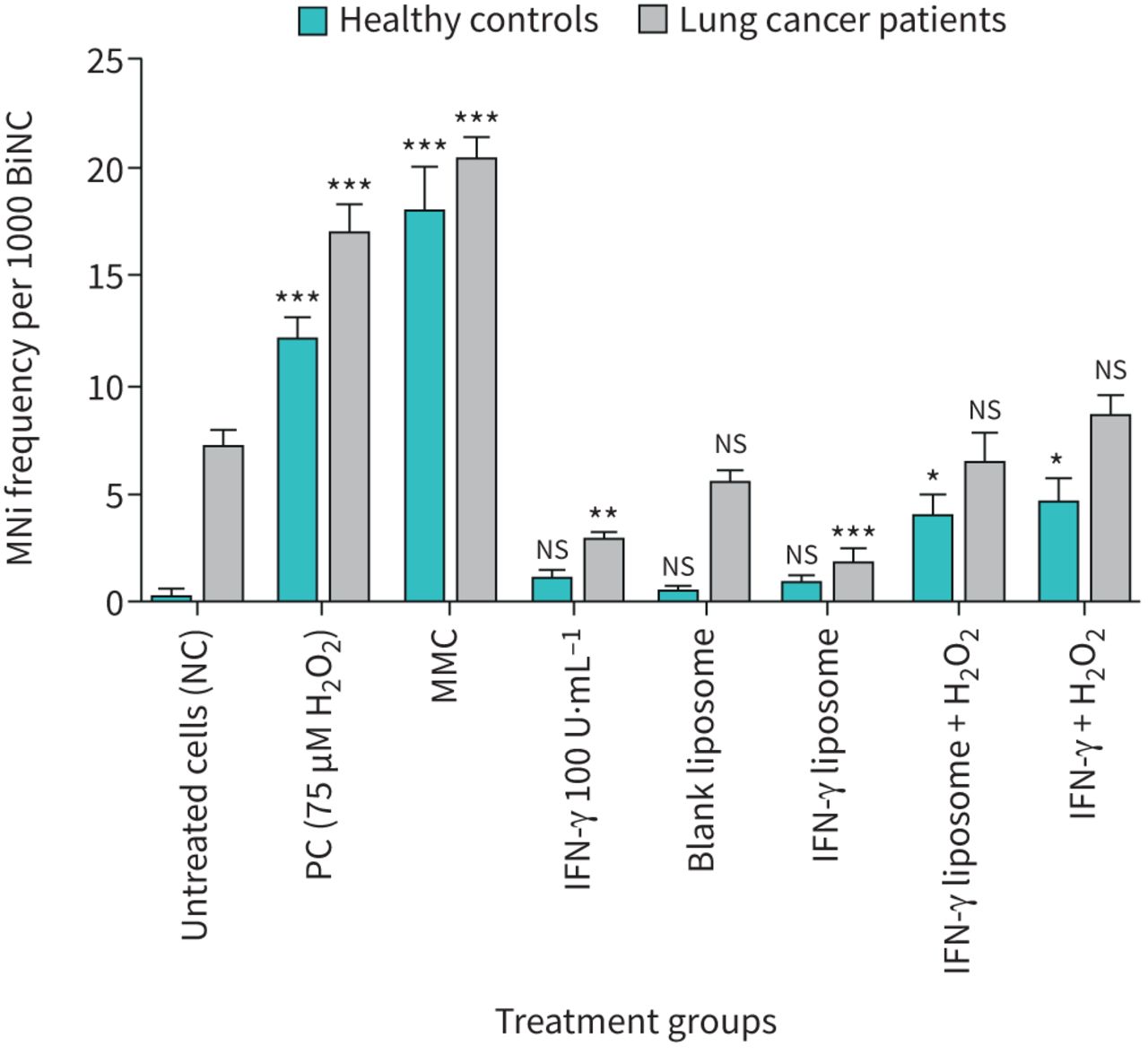
The lymphocytes from healthy individuals and lung cancer patients were treated with 75 μM H2O2, 0.4 μM MMC, 100 U·mL−1 of IFN-γ bulk, IFN-γ liposome and 100 U·mL−1 of IFN-γ bulk, IFN-γ liposome with 75 μM H2O2. They were then assessed for MNi induction using the CBMN assay, as presented in figure 4. In contrast, lymphocytes from healthy individuals treated with 100 U·mL−1 of IFN-γ bulk and IFN-γ liposome showed no effect on MNi frequency compared to untreated cells. The MMC and 75 μM H2O2 displayed a significant increase in the MNis of lymphocytes (p<0.001), but 100 U·mL−1 of IFN-γ bulk and IFN-γ liposome co-supplemented with H2O2 showed a significant decrease in MNis of lymphocytes (p<0.05) as in figure 4. It was evident that the 100 U·mL−1 of IFN-γ bulk and IFN-γ liposome showed a significant reduction in the number of MNis in lung cancer patients, but the reduction by IFN-γ liposome was greater (p<0.001) compared to untreated cells. Furthermore, the MMC and 75 μM H2O2 showed a significant increase (p<0.001) in the MNi number on lymphocytes compared to the untreated cells.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comparison of MNi frequencies between healthy individuals and lung cancer patients, after treatment with 75 μM H2O2, 0.4 μM Mitomycin C (MMC), 100 U·mL−1 of interferon (IFN)-γ bulk, IFN-γ liposome and 100 U·mL−1 of IFN-γ bulk, IFN-γ liposome in the presence of 75 μM H2O2. n=5 in each group. BiNC: binucleated cells; NC: nucleated cells; PC: positive control; ns: nonsignificant. *: p<0.05; **: p<0.01; ***: p<0.001.
There are numerous cytological scoring parameters, including biomarkers of cell mitotic division such as mononucleated cells (MoNCs), binucleated cells (BiNCs) and multinucleated cells (MultiNCs). From these parameter values, the NDI was calculated for healthy individuals and lung cancer patients, respectively. The mean values for the NDI for all treatment groups were within the normal range, as shown in table 2. Table 2 also displays the chromosomal instability parameters in the form of the mean of NPBs, NBUDs per 1000 BiNC and MNi frequency in MoNC. The table clearly shows that the frequency of MNi in BiNC of untreated cells is higher than in lymphocytes treated with IFN-γ liposome on lung cancer patients.
The mean of different parameters for chromosomal damage in healthy individuals and lung cancer patients lymphocyte using the cytokinesis block micronucleus assay
Discussion
This study investigated the efficacy of IFN-γ and IFN-γ liposome in human lymphocytes originating from groups of 20 healthy individuals and 20 lung cancer patients, using the Comet assay and five individuals in each group for the CBMN assay. Different in vitro and in vivo studies have shown the promising applications of IFN-γ in the field of health as a treatment displaying pleiotropic immunomodulatory, antiviral, antimicrobial, antineoplastic, proinflammatroy activities [12]. Furthermore, the potent inhibitory effects of IFN-γ in various tumour models, such as bladder carcinoma, colorectal cancer, ovarian cancer, and adult T cell leukaemia, human pancreatic carcinoma cells, and NSCLC have been evidenced [11, 17].
Research has demonstrated that IFN-γ can induce apoptosis in human glioblastoma T98G and U87MG cells [41], and inhibited the proliferation of four pancreatic cancer cell lines in vitro [42]. Other studies indicated that IFN-γ was responsible for apoptosis in ovarian cancer cells in vivo and in vitro [43]. By contrast, none of the previous studies has focused on the effect of IFN-γ on DNA damage in human lymphocytes from lung cancer patients through the use of the Comet and CBMN assay.
The obtained results showed that the treated lymphocytes from healthy individuals with IFN-γ bulk and IFN-γ liposome did not induce significant DNA damage when compared to the untreated cells. However, when the same concentration of IFN-γ in both forms was used on lung cancer patients' lymphocytes, DNA damage was reduced compared to the untreated cells. Still, the reduction of DNA damage that occurred by the liposome form of IFN-γ was greater than from the IFN-γ bulk (figures 2 and 3). This could possibly be due to the biocompatibility and enhanced cellular interaction of liposomes compared to their larger particles [44]. The Comet assay results showed that IFN-γ bulk and the liposome form have a protective effect against H2O2, which induces damage in DNA due to oxidative stress in healthy individuals and lung cancer patients’ lymphocytes. DNA damage significantly decreased compared to the PC. The effective results of IFN-γ liposome against H2O2-induced damage could be due to the ability of liposome in preventing cell membrane alterations rather than its direct interaction with H2O2 [45].
The micronucleus assay was used to investigate the influence of IFN-γ bulk and liposome on the lymphocytes from healthy individuals and lung cancer patients at the chromosomal level. The CBMN assay has been a critical test in detection of the genotoxicity of different compounds by measuring MNi and other chromosomal abnormalities such as NPBs, a biomarker of dicentric chromosomes, and by measuring NBUDs, which are a biomarker of gene amplification [46]. MNis are cytoplasmic bodies generated as a consequence of disorders during cell cycle division. Subsequently, when the centric fragments or whole chromosomes are incapable of travelling to opposite poles during anaphase, the nuclear envelope forms around the lagging chromosomes and fragments, which gradually take on the morphology of an interphase nucleus, which is smaller than the main nuclei in the cell [36, 47]. In this assay, the dividing cell cytokinesis is inhibited with cytochalasin B (Cyto B) [48].
In this study, amongst the different types of cells counted, specific consideration was made for the presence of MNi in the treatment groups because MNi are indicators of chromosomal breakage, loss, rearrangement, necrosis and apoptosis [35]. Furthermore, MNi in BiNC only showed the damage after treatment, which decreased the probability of scoring the pre-existing damage [49]. Mitomycin C (MMC) is recognised as an antitumour antibiotic, clastogenic and genotoxic compound. Therefore, MMC served as a PC in this study. MMC induced MNi production in BiNCs more than mononucleated cells (table 2). The percentage of BiNC after scoring 1000 cells was within the normal range in lymphocyte cultures for each individual. On the other hand, the percentage of MultiNC in cell culture was low, thus elucidating that cytochalasin B inhibited cell division after one cell cycle. Meanwhile, the NDI values were normal for all experiments (table 2).
From this assay, the frequency of MNi in the lymphocytes from healthy individuals and lung cancer patients decreased when treated with IFN-γ and IFN-γ liposome compared to the untreated cells (NC) for each group. In contrast, the IFN-γ liposome showed a greater reduction in MNi frequency in lymphocytes. H2O2 was used in this assay and functioned as expected, inducing MNi production on the lymphocytes from both healthy individuals and lung cancer patients. The combination of IFN-γ bulk and IFN-γ liposome with H2O2 reduced the frequency of MNis compared to H2O2 in patients’ lymphocytes. This study's most important finding was that IFN-γ liposome had exhibited higher DNA damage reduction effects at 100 U·mL−1 concentration than IFN-γ bulk.
The Comet assay data showed that IFN-γ in bulk and liposome form reduced DNA damage in the lymphocytes from healthy individuals and lung cancer patients compared to the NC.
Our results were consistent with a previous study where IFN-γ induced early stimulation of PARP (the DNA repair enzyme) when the cells were exposed to a proapoptotic agent [50]. Furthermore, DNA damage was decreased when the IFN-γ bulk and liposome were treated with H2O2 in the lymphocytes from healthy individuals and lung cancer patients compared to the PC. This denotes the ability of IFN-γ to protect against DNA damage in lymphocytes from oxidative stress. The CBMN assay followed a similar pattern to the Comet assay results. It also showed a reduction in MNi frequency in the lymphocytes, thus implying the potential facilitation of DNA repair.
In general, IFN-γ liposome (100 U·mL−1) has shown better protective effects than its larger particle counterpart, bulk, at the same concentrations of 100 U·mL−1 each. This could perhaps be due to the enhanced solubility of liposome-coated IFN-γ in an aqueous medium stabilising various therapeutic agents such as proteins and nucleotides. Their size, surface charge and composition are favourable conditions for their effective results [51]. Many hydrophilic and hydrophobic biomolecules can be encapsulated in liposomes and protected from interactions with the external environment. Liposomes inhibit the metabolism of the drug before reaching the target cells plus protect the drug's interaction with healthy cells, enhancing the therapeutic index of the drug. It has been well documented that the liposome form of any therapeutic drug shows enhanced pharmacokinetic properties and biodistribution than its free form. The morphology of liposomes is similar to that of the cellular membranes, making them an ideal drug carrier system [44].
In conclusion, the data obtained from the Comet and micronucleus assays indicated that IFN-γ in both forms might potentially be effective for lung cancer patients. Overall data from the present study proposes that IFN-γ could protect and defend against lung cancer through cell cycle arrest of cancer cells and repair mechanisms and liposomes can potentially be used as an alternative, better drug delivery system in various conditions.
Acknowledgement
The authors would like to thank the clinical staff of Bradford Royal Infirmary and St Luke's Hospital for their assistance in this study.
Footnotes
This article has been revised according to the author correction published in ERJ Open Res 2021; 7: 50555-2020 [https://doi.org/10.1183/23120541.50555-2020].
Conflict of interest: M. Alhawmdeh has nothing to disclose.
Conflict of interest: M. Isreb has nothing to disclose.
Conflict of interest: A. Aziz has nothing to disclose.
Conflict of interest: B.K. Jacob has nothing to disclose.
Conflict of interest: D. Anderson has nothing to disclose.
Conflict of interest: M. Najafzadeh has nothing to disclose.
Support Statement: The sponsorship by the Mutah University of a PhD studentship to Maysa Alhawamdeh is gratefully acknowledged. The sponsor played no part in the conduct of the work or the writing of the manuscript. Funding information for this article has been deposited with the Crossref Funder Registry.
- Received August 5, 2020.
- Accepted March 23, 2021.
- Copyright ©The authors 2021
This version is distributed under the terms of the Creative Commons Attribution Non-Commercial Licence 4.0. For commercial reproduction rights and permissions contact permissions{at}ersnet.org
References










