





Abstract
Background Individuals with sarcoidosis are at higher risk for infection owing to underlying disease pathogenesis and need for immunosuppressive treatment. Current knowledge as to how subjects with sarcoidosis respond to different forms of vaccination is limited. We examined quantitative and functional antibody response to COVID-19 vaccination in infection-naive subjects with and without sarcoidosis.
Methods Our prospective cohort study recruited 14 subjects with biopsy-proven sarcoidosis and 27 age–sex matched controls who underwent a two-shot series of the BNT162b2 mRNA vaccine at the University of Illinois at Chicago. Baseline, 4-week and 6-month trimer spike protein IgG and neutralising antibody (nAb) titres were assessed. Correlation and multivariate regression analysis was conducted.
Results Sarcoidosis subjects had a significant increase in short-term antibody production to a level comparable to controls; however, IgG titres significantly declined back to baseline levels by 6 months. Corresponding neutralising assays revealed robust nAb titres in sarcoidosis subjects that persisted at 6 months. A significant and strong correlation between IgG and nAb titres across all time points was observed in the control group. However within the sarcoidosis group, a significant but weak correlation between antibody levels was found. Overall, IgG levels were poor predictors of nAb titres at short- or long-term time points.
Conclusions Sarcoidosis subjects exhibit nAb induced by the BNT162b2 mRNA SARS-CoV-2 vaccine at levels comparable to controls that persists at 6 months indicating conferred immunity. Trimer IgG levels are poor predictors of nAb in subjects with sarcoidosis. Studies of further antibody immunoglobulins and subtypes warrant investigation.
Abstract
While subjects with sarcoidosis have a diminished quantitative antibody (anti-trimer) response to the BNT162b2 mRNA #COVID19 vaccine, their functional neutralising antibody response is comparable to controls, indicating conferred immunity https://bit.ly/3TtqUQZ
Introduction
Since the start of the COVID-19 pandemic, the development of effective treatments to diminish COVID-19 disease severity has been an international priority. Vaccines were developed at record speed and offer a life-changing opportunity for disease mitigation and prevention. Initial studies demonstrated the mRNA-based COVID-19 vaccines, BNT162b2 and mRNA-1273, were efficacious in preventing up to 95% and 94.1% of COVID-19 disease in recipients, respectively [1, 2]. However, vaccine response in vulnerable populations remains ill defined.
Sarcoidosis is a multisystem disease of unknown aetiology characterised by granulomatous inflammation and subsequent organ dysfunction. This inflammation is believed to stem from maladaptive immune responses, resulting from chronic immune stimulation with subsequent risk of lymphocyte anergy, exhaustion and depletion [3–5]. Data supports sarcoidosis subjects as having increased risk of infection [6, 7], though the overall risk of SARS-CoV-2 is unclear [8]. Beyond immune susceptibility conferred by underlying disease pathology, nearly one-fourth of all sarcoidosis patients require treatment with immunosuppressive agents, which further contributes to infectious risk [9]. Primary infection prevention with vaccination in this population is therefore of great importance.
Literature regarding how individuals with sarcoidosis respond to vaccines is limited and indicates varying responses. A study of tetanus vaccination in sarcoidosis patients found 50% had an insufficient increase in antibody titres regardless of sarcoid disease state, stage or duration, and independent of treatment [10]. A separate study of a three-dose series of the hepatitis B vaccine found that none of the 16 sarcoidosis subjects had detectable antibody levels at 1 month follow-up [11]. In contrast, a study of the 2008–2009 trivalent influenza vaccine showed sarcoidosis and control subjects had a comparable serological response [12]. In addition, existing literature explores quantitative assessment of antibody response through measurement of immunoglobulin titres; however these titres may not ensure conferred protective immunity. While developed antibodies may target any viral epitope, neutralising antibodies (nAb) bind to the virus in such a way that it inhibits cell entry and/or viral replication therefore blocking infection from propagating [13]. Post-vaccination nAb assays provide insight into the functional protection allocated by the vaccine, and to our knowledge there are no current studies evaluating nAb in sarcoidosis.
With regard to immunosuppression, data assessing the effect of immunosuppressive medication on vaccine efficacy in sarcoidosis are limited, and recommendations are extrapolated from studies of other immune-related disorders. Use of various immunosuppressive medications is associated with decreased antibody response to multiple types of vaccinations, including the mRNA COVID vaccines [14–17]. Regardless of a potential insufficient response, vaccination is strongly recommended in sarcoidosis to protect against various community-acquired infections to include COVID-19 [18, 19].
We postulate that subjects with sarcoidosis will have a deficient immune response to COVID-19 vaccination. This study aims to characterise the antibody response to COVID-19 vaccination in subjects with and without sarcoidosis through quantitative assessment of binding antibodies and correlation to functional assessment of nAb. Our findings may direct vaccination guidelines, inform the need for further booster vaccines and extrapolate further information about the immune dysregulation underlying sarcoidosis pathology.
Methods
Study population and sample acquisition
Study approval was obtained through the University of Illinois at Chicago (UIC) IRB Ethics Review Committee, Approval #2018-1038.
Subjects with biopsy-proven sarcoidosis, diagnosed in accordance with ATS/ERS/WASOG criteria [20], and who were undergoing vaccination with the BNT162b2 mRNA COVID-19 vaccination were recruited. All subjects were older than 18 years of age and receive their sarcoidosis care in the Bernie Mac Sarcoidosis Translational Advanced Research (STAR) Center at UIC. Demographic and clinical data were extracted from the electronic medical record and included sex, race, age, body mass index (BMI) and sarcoidosis organ involvement, as well as treatment with immunosuppressive therapy (systemic steroids and/or disease modifying anti-sarcoid drugs (DMASDs)). Peripheral lymphocyte counts in the preceding 6 months were also recorded. Age and sex-matched self-reported-immunocompetent control subjects consisted of University of Illinois Hospital employees who were undergoing vaccination at UIC. Demographic and clinical data were collected using a questionnaire and included race, sex, age, height and weight, medication use, and existing medical problems. Any subject who self-reported a personal history of COVID-19 infection was excluded.
Blood samples were collected at baseline (just prior to first vaccine dose, timepoint V1D0), 4 weeks (i.e. 7 days after the booster dose, timepoint V2D7) and 6 months after the first vaccine dose (time point M6). Serum was extracted within 2 h of sample collection and stored at −80°C.
Anti-spike (trimer) IgG titer quantification
The Human SARS-CoV-2 Spike Trimer IgG ELISA Kit (Catalog #BMS2325, ThermoFisher Invitrogen, Waltham, MA, USA) was used to quantitate serum IgG levels of each subject at each timepoint, per the manufacturer's protocol. All samples were diluted 1:100 and assayed in duplicate with a series of 2-fold serial dilutions (1:150 000 units·mL−1 to 1:2340 units·mL−1) of the standard control for relative quantification. Absorbance at 450 nm was quantified using a Spark® multimode microplate reader (Tecan, Mannedorf, Switzerland). Samples that produced signals greater than the upper limit of the standard curve were reassayed at 1:1000 dilution. IgG concentration was calculated by fitting 5-parameter logistic curves to the standard controls and reported as binding antibody units per millilitre (BAU·mL−1). The mean concentration of duplicates was utilised for analysis. Inter-assay variability was addressed utilising the ELISAtools package in R (version 4.0.4, Boston University School of Medicine, Boston, MA, USA) to account for batch-effect [21].
Antibody neutralisation assays
Neutralisation activity against SARS-2-CoV was measured in a single-round-of-infection assay using pseudotyped viruses [22]. Briefly, 293T angiotensin-converting enzyme (ACE) cells were infected with modified vesicular stomatitis virus (VSV), which lacks a gene vital for VSV replication and instead carries a firefly luciferase reporter gene that allows for chemiluminescence. The cells are also transfected with plasmids encoding full length SARS-CoV-2 spike (S) protein, which is a surface protein responsible for binding the host cell receptor, ACE2, to mediate viral entry during SARS-CoV-2 infection (SARS-CoV-2 Spike-pseudotyped lentiviral particle kit, Catalog #NR-53816, BEI Resources, Manassas, VA, USA). The pseudotyped virus therefore contains the SARS-CoV-2 S protein to simulate viral entry and VSV, which provides the structural genes for viral packaging without the ability to replicate. The 50% tissue culture infectious dose (TCID50) of the pseudotyped virus, which indicates the amount of virus required to quash 50% of the inoculated cells, was calculated according to the Reed–Muench method.
Serial dilutions of each subject's serum were incubated for 1 h at 37°C with 1000 TCID50/mL of the pseudotyped virus (virus plus antibody) then added to monolayers of ACE2-overexpressing 293T cells in quadruplicate on a 96-well plate. Controls consisted of pseudotyped virus and 293T cells without added serum sample (virus-only). The plate was incubated for 65–72 h in the cell culture incubator at 37°C and 5% (vol/vol) CO2 after which 50 µL of luciferase substrate was added to stimulate chemiluminescence. The amount of chemiluminescence, determined by a plate reader, directly correlates with the amount of pseudotyped virus that has entered and “infected” the cells. The amount of nAbs, which inhibit viral entry into the 293T cell, is therefore inversely correlated with the chemiluminescence signal intensity. Neutralising antibody titres are reported as the 50% inhibitory dilution (ID50), calculated using the Reed–Muench method, which refers to the dilution fold required to achieve 50% neutralisation [22, 23]. The initial serum dilution was 1:25; therefore 25 is the cut-off for antibody detection. Higher ID50 correlates to increased potency of nAb within the serum sample.
Statistical analysis
Demographic data was tested for significance between groups utilising the Mann–Whitney U-test (MWU) for nonparametric continuous data, or Fisher's exact-test for categorical data, as appropriate. Statistical differences between groups of time from vaccination to sample collection were assessed using the t-test. The primary outcome measures were absolute post-vaccination trimer IgG titres and nAb titres at both V2D7 and M6, as neither assay has a validated cut-off indicating test positivity. Antibody titres for all time points were log transformed and z-scores were calculated to identify outliers (z-score ≥2.5). Titres were tested for significance using the MWU test separately for each time point. Subgroup analysis was performed to assess the role of immunosuppressive therapy, with differences tested using the Kruskal–Wallis test with Dunn's post hoc test and Benjamini–Hochberg correction for multiple comparisons. Univariate correlation analysis was performed by calculating Pearson's coefficients for log transformed antibody titres. Multivariate regression models were then constructed separately for trimer IgG and nAb to determine the relative effect of significant baseline variables on short-term (V2D7) and long-term (M6) results. p-values <0.05 were considered significant. All analyses were performed in R version 4.0.4 (www.R-project.org/). Kruskal–Wallis and Dunn tests were implemented using the Dunn.test package. T-testing, MWU and Fisher's exact-testing was implemented using the stats package. Pearson coefficients were calculated using the corr.test function of the psych package.
Results
Demographics
14 sarcoidosis subjects and 27 control subjects were recruited. Group characteristics are highlighted in table 1. Nearly all subjects received the BNT162b2 mRNA COVID-19 vaccine and booster at the recommended time interval of 21 days (mean±SD 21.15±0.57 days, range 20–24 days). All subjects had blood samples collected just prior to the first vaccine dose administration (V1D0) and 7 days after the booster dose at time point V2D7 (mean±SD 6.91±0.28 days, range 6–7 days). All recruited subjects had available samples for trimer IgG analysis at V1D0 and V2D7; of whom 17 control samples and all 14 sarcoidosis samples were included in nAb analysis. Owing to attrition, 22 control subjects and 11 sarcoidosis subjects were available for blood samples at the 6-month time point (mean±SD 184.8±12.2 days from first vaccine, range 170–214 days) and included in trimer IgG analysis at M6, of whom 11 control subjects and all 11 sarcoidosis subjects were included in nAb analysis.
Demographics of control and sarcoidosis groups
The sarcoidosis group comprised 13 subjects with pulmonary manifestations and six with extrapulmonary involvement. Six subjects were not on any treatment, and eight were treated with immunosuppressive therapy. Specifics regarding sarcoidosis phenotypes and treatment regimens for each subject are described in table 2. Absolute peripheral lymphocyte values were available for 12 sarcoidosis subjects and had a group median of 1.8×109 cells·L−1. Four subjects had medication titration within this 6-month pre-vaccination time interval.
Clinical details regarding sarcoidosis chronicity, organ involvement and treatment for each of the 14 subjects in the sarcoidosis group
Anti-spike protein trimer IgG titer and neutralising antibody analysis
Trimer IgG titres for each group across all three time points are illustrated in figure 1. There were no outliers detected at either time point. While sarcoidosis subjects had a higher median baseline IgG titre than the control group (MWU p-value <0.001), both groups demonstrated a significant increase in IgG titres at V2D7 compared to their respective baselines (sarcoidosis: MWU p-value <0.001; control: MWU p-value <0.001) with comparable titres at V2D7 between groups (MWU p-value = 0.3680). IgG titres in both groups significantly decreased at the M6 time point from their respective V2D7 titres (sarcoidosis: MWU p-value <0.001; control: MWU p-value <0.001); however, M6 IgG titres in the sarcoidosis group fell to levels comparable to sarcoidosis V1D0 titres (MWU p-value = 0.9786) and were significantly less than M6 control IgG titres (MWU p-value = 0.0237). M6 IgG titres in the control group remained significantly higher than baseline values (MWU p-value <0.001). Overall, this trend indicates a robust initial IgG response in both groups that diminishes over time, returning to baseline in the sarcoidosis group and raising the concern for more transient antibody protection.

Trimer IgG titres for control and sarcoidosis groups are shown: a) log transformed titres for comparison between time points for each group; and b) log transformed titres for comparison between groups at each time point. NS: p>0.05; *: p≤0.05; ***: p≤0.001. NS: nonsignificant.
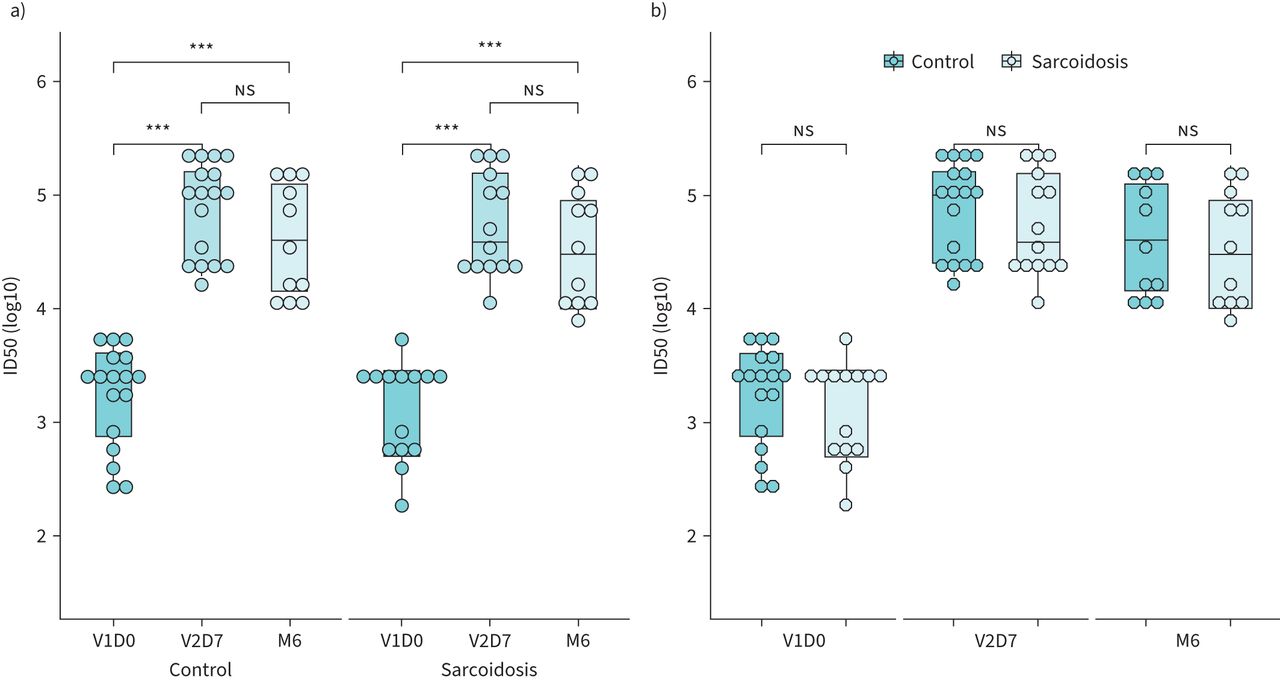
Functional nAb assays were performed to better determine protection conferred from vaccination. The ID50 for each group across all three time points (V1D0, V2D7 and M6) is illustrated in figure 2. In contrast to significantly elevated IgG in the sarcoidosis group at baseline, there was no significant difference in ID50 between groups at the V1D0 time point (MWU p-value = 0.5879). Both control and sarcoidosis groups had a significant increase in nAb titres from baseline to V2D7 suggesting robust nAb formation after 1st and 2nd vaccination doses (sarcoidosis: MWU p-value <0.001; controls: MWU p-value <0.001), similar to what was observed for IgG trends. However, median ID50 for both groups at M6 were not significantly changed from their respective V2D7 values (sarcoidosis: MWU p-value = 0.2250; controls: MWU p-value=0.0894) and remained significantly higher than baseline (sarcoidosis: MWU p-value <0.001; controls: MWU p-value <0.001) suggesting persistent immunity. Finally, median ID50 values for sarcoidosis subjects were comparable to those of controls at all time points (V1D0 MWU p-value = 0.5879; V2D7 MWU p-value = 0.5740; M6 MWU p-value = 0.7409) indicating similar levels of nAb present.

Neutralising titres (50% inhibitory dilution (ID50)) for control and sarcoidosis groups are shown: a) log transformed titres for comparison between time points for each group and b) log transformed titres for comparison between groups at each time point. NS: p>0.05; ***: p≤0.001.
Trends in IgG and nAb were further evaluated across sarcoidosis treatment groups (figure 3). Pairwise comparisons using Dunn's test indicated sarcoidosis subjects both on and off immunosuppression had comparable V2D7 IgG titres to controls (immunosuppression versus control Benjamini–Hochberg (BH) adjusted p-value = 0.3197, no treatment versus control BH adjusted p-value = 0.4514) as well as comparable nAb titres to controls (immunosuppression versus control BH adjusted p-value = 0.4904, no treatment versus control BH adjusted p-value = 0.9333) indicating a robust initial antibody response regardless of immunosuppression use. At M6, sarcoidosis subjects on immunosuppression had significantly decreased IgG titres compared to controls (BH adjusted p-value = 0.0162); however nAb titres remained comparable (controls BH adjusted p-value = 0.3688) suggesting preserved protection.
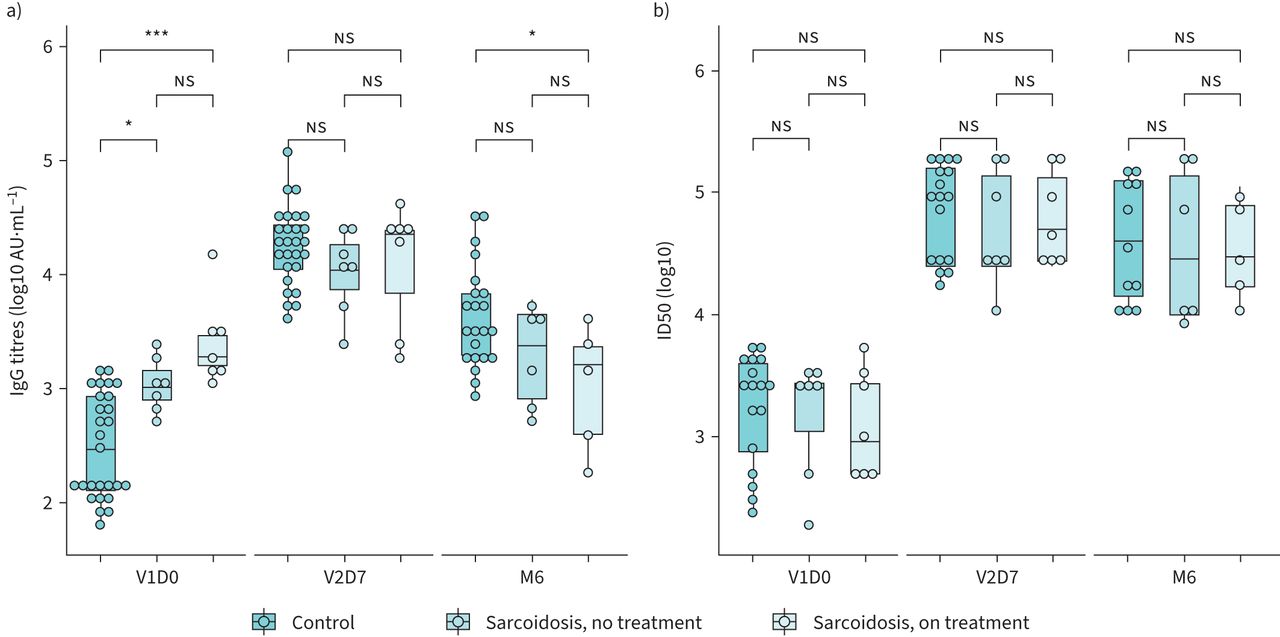
a) log transformed trimer spike protein IgG titres are shown. Sarcoidosis subjects not on treatment had significantly higher V1D0 titres than controls and comparable titres at V2D7. M6 IgG titres were significantly lower in the sarcoidosis group than controls. b) log transformed neutralising titres (50% inhibitory dilution (ID50)). Values were comparable across all groups at each time point. NS: p>0.05; *: p≤0.05; ***: p≤0.001.
Regression analysis
Given variation in trimer IgG and nAb trends, a univariate linear regression model was constructed to characterise the relationship between IgG and nAb titres across all time points. Correlation coefficients for control and sarcoidosis group are shown in figure 4. IgG titres were significantly and directly associated with nAb titres in both groups, with a strong correlation for the control group (R = 0.7715, p-value <0.001) and a weak correlation for sarcoidosis group (R=0.3905, p-value = 0.0140). IgG titres in the sarcoidosis group were overall determined to be less predictive of nAb (R2 = 0.1525) than in the control group (R2 = 0.5952). With such low variance explained by IgG titres in the sarcoidosis group, a multivariate linear regression model was subsequently constructed to delineate which, if any, variables independently predict short-term (V2D7) and long-term (M6) nAb titres. Regression models for each outcome are shown in figure 5. The overall regression was not statistically significant for either group. Additional regression models for M6 for each group were constructed with the addition of V2D7 and M6 IgG titres, also with interdependencies accounted for, and yielded similar results. None of the additional independent variables were significantly predictive of nAb though analysis may be underpowered to detect significance.

Univariate linear regression analysis illustrating the relationship between log transformed trimer IgG titres and log transformed 50% inhibitory dilution (ID50) across all time points. a) Control group showing a significant and strong correlation; b) sarcoidosis group showing a significant yet weak correlation.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Multivariate regression analysis to assess independent predictors of 50% inhibitory dilution (ID50) by group (top row: controls; bottom row: sarcoidosis) and outcome time points (left column: V2D7; right column: M6). Axes are log transformed. a) V2D7 ID50 for control group. Model: Log10 V2D7 ID50 ∼ Log10 V2D7 Trimer IgG * Log10 V1D0 Trimer IgG + Race + BMI. b) M6 ID50 for the control group. Model: Log10 V2D7 ID50 ∼ Log10 V2D7 Trimer IgG * Log10 V1D0 Trimer IgG + Log10 V2D7 Trimer IgG * Log10 M6 V2D7 IgG + Race + BMI. c) V2D7 ID50 for the sarcoidosis group. Model: Log10 V2D7 ID50 ∼ Log10 V2D7 Trimer IgG * Log10 V1D0 Trimer IgG + Race + BMI + Treatment Group. d) M6 ID50 for the sarcoidosis group. Log10 V2D7 ID50 ∼ Log10 V2D7 Trimer IgG * Log10 V1D0 Trimer IgG + Log10 V2D7 Trimer IgG * Log10 M6 V2D7 IgG + Race + BMI + Treatment Group. The overall regression was not statistically significant for either group at both time points.
Discussion
We present a single centre analysis of the quantitative and qualitative antibody response to vaccination with the BNT162b2 mRNA COVID-19 vaccine in infection naive subjects with and without sarcoidosis. Our data indicate that sarcoidosis subjects mount a robust initial trimer IgG antibody response to vaccination with subsequent quantitative decline by 6 months, driven by those on immunosuppression. Despite the decline in binding antibodies, sarcoidosis subjects develop and maintain functional immunity regardless of immunosuppressive treatment. With this discrepancy between IgG and nAb titres, it is not surprising that IgG antibodies weakly correlated with nAb and were not significantly predictive of nAb titres at any timepoint. While this study is of a single vaccine type, it sheds light on the clinical protection vaccination provides individuals with sarcoidosis despite IgG titres that may suggest otherwise.
The SARS-CoV-2 is an enveloped virus with numerous structural proteins vital for the viral life cycle. The S protein, which is composed of a S1 subunit, S2 subunit, N-terminal domain and receptor-binding domain (RBD), is responsible for viral entry into the host cell specifically through binding of the RBD. Antibodies to SARS-CoV-2 consist of IgM, IgG and/or IgA antibodies and may target any subdomain of the S protein [24–27]. Upon viral binding to ACE2, conformational changes of surface glycoproteins result in the formation of an S protein trimer [28], which is the target of the IgG antibodies quantified in this study. Despite a wide array of antibodies produced, nAb confer protection by preventing viral entry, receptor-mediated infection and infection propagation. Numerous studies support a strong correlation between various antibody titres and neutralisation; however, the strength of correlation may vary with different tested antibody targets as well as with time from SARS-CoV-2 exposure [29–31]. Our findings reveal a strong correlation in controls between trimeric anti-spike IgG levels and neutralisation, which aligns with a similar study of this antibody type in healthy individuals after BNT162b2 vaccine [32]. The sarcoidosis group, however, had a weak correlation between IgG levels and neutralisation which was otherwise not explained by group differences. Neutralisation in this group is therefore likely explained by the presence of non-IgG antibodies or antibodies to other spike protein subdomains not measured in our study, though further analyses are needed to assess this.
Sarcoidosis is a T-cell-mediated disease characterised by local CD4+ T-lymphocyte inflammation and peripheral lymphocyte depletion in severe or active disease, as well as anergy and exhaustion in progressive disease [4, 5, 33, 34]. Defects within humoral immunity have also been described and include evidence of B-cell hyperactivation, autoantibody production and decreased circulating memory B-cells, as well as previously mentioned impaired serologic responses to tetanus and hepatitis vaccines [10, 11, 20, 35–38]. Despite these defects, vaccine-induced development and persistence of neutralisation antibodies in sarcoidosis subjects is a particularly important and reassuring finding. However, it is worth noting the median peripheral absolute lymphocyte count (1.8×109 cells·L−1) in our cohort is greater than previously described thresholds of significant sarcoidosis-related lymphopenia [34]. This suggests lymphopenia is unlikely to be a significant disease manifestation in this cohort and therefore may explain the preserved nAb activity. Treatment was also uptitrated in four sarcoidosis individuals; despite this, neutralisation was seemingly unaffected.
Limitations of this study include heterogeneity within the sarcoidosis group, limited sample size and attrition rate at 6 months. Data regarding lymphocyte subsets in the sarcoidosis group were largely unavailable and may have allowed further interpretation of the immunity stimulated by vaccination if obtained concurrently. Only IgG antibody was quantified, and despite strong correlation to nAb among control subjects, further assessment of preserved immunity in sarcoidosis was limited. Future studies in the field should focus on inclusion of specific sarcoidosis phenotypes as well as direct assessment of cellular and humoral activity. Furthermore, baseline elevation of the trimer IgG antibody in the sarcoidosis group raised the question of nonspecific IgG binding from underlying hypergammaglobulinaemia [39] or prior community coronavirus exposure with cross-reactivity [40]. Undiagnosed low-level exposure or infection in the sarcoidosis group is also considered as COVID-19 history was self-reported, though the similar levels of baseline neutralising antibodies does not corroborate this. Lastly, only one mRNA COVID-19 vaccine was studied; thus conclusions should be cautiously applied to other COVID-19 vaccines.
Despite these limitations, we conclude that trimer IgG levels in sarcoidosis subjects are poor predictors of nAb, which are an important mechanism in preventing infection. While further analysis is needed to determine clinical outcomes from vaccination in this vulnerable population, particularly among those on immunosuppression, knowledge gained from our study suggests that vaccination may provide at least partial protection from COVID-19 infection in sarcoidosis. Additional studies of immune response stimulated by the BNT162b2 vaccine, which induces robust cellular and humoral immunity [41], may offer mechanistic insights into the pathogenesis of sarcoidosis.
Supplementary material
Supplementary Material
Please note: supplementary material is not edited by the Editorial Office, and is uploaded as it has been supplied by the author.
Supplementary material 00025-2022.SUPPLEMENT
Acknowledgements
The authors acknowledge the contributions of Sunghyun Hwang (Department of Medicine, University of Illinois at Chicago (UIC)), Rhea Goel (Department of Medicine, UIC), Benjamin A. Turturice (Department of Medicine, Department of Microbiology and Immunology, UIC), Cody Schott (Department of Medicine, Department of Microbiology and Immunology, UIC), Montserrat Hernandez (Department of Medicine, UIC), Yang Chen (Department of Medicine, Department of Microbiology and Immunology, UIC), Julianne Jorgensen (Department of Medicine, Department of Bioengineering, UIC), Wangfei Wang (Department of Medicine, Department of Bioengineering, UIC) and Mladen Rasic (Department of Medicine, Department of Bioengineering, UIC).
Footnotes
Provenance: Submitted article, peer reviewed.
Author contributions: Conception and study design: C.L. Vagts, Y. Chang, C. Ascoli, D.L. Perkins and P.W. Finn. Participant recruitment, sample collection and sample processing: C.L. Vagts, Y. Chang, J.M. Lee, K. Huang, C. Ascoli, Y. Huang, N. Sarup, S.R. Warpecha, R.A. Cherian, R. Edafetanure-Ibeh and N.J. Sweiss. Experiments: C.L. Vagts, Y. Chang, J.M. Lee, N. Sarup, R Cherian, M. Amin, T. Sultana, M. Ghassemi and R. Novak. Analysis and interpretation: C.L. Vagts, Y. Chang, C. Ascoli and N. Sweiss. Drafting the manuscript for important intellectual content: C.L. Vagts, Y. Chang, K. Huang, Y. Huang, C. Ascoli, N.J. Sweiss and P.W. Finn. C.L. Vagts acts as the guarantor of the paper.
Conflict of interest: Richard M. Novak reports the following relationships outside the submitted work: grants or contracts received from Janssen; consulting fees received from Gilead and Viiv. The remaining authors have nothing to disclose.
Support statement: Support for this study was provided by NIH grant numbers T32 HL144909, R01 HL138628, R01 HL138628-01A1S1, F30 HL151182 and F30 HD102093. Funding information for this article has been deposited with the Crossref Funder Registry.
- Received January 13, 2022.
- Accepted August 19, 2022.
- Copyright ©The authors 2023
This version is distributed under the terms of the Creative Commons Attribution Non-Commercial Licence 4.0. For commercial reproduction rights and permissions contact permissions{at}ersnet.org
References










