





Abstract
Background COPD patients are more susceptible to viral respiratory infections and their sequelae, and have intrinsically weaker immune responses to vaccinations against influenza and other pathogens. Prime-boost, double-dose immunisation has been suggested as a general strategy to overcome weak humoral response to vaccines, such as seasonal influenza vaccination, in susceptible populations with weak immunity. However, this strategy, which may also provide fundamental insights into the nature of weakened immunity, has not been formally studied in COPD.
Methods We conducted an open-label study of seasonal influenza vaccination in 33 vaccine-experienced COPD patients recruited from established cohorts (mean age 70 (95% CI 66.9–73.2) years; mean forced expiratory volume in 1 s/forced vital capacity ratio 53.4% (95% CI 48.0–58.8%)). Patients received two sequential standard doses of the 2018 quadrivalent influenza vaccine (15 μg haemagglutinin per strain) in a prime-boost schedule 28 days apart. We measured strain-specific antibody titres, an accepted surrogate of likely efficacy, and induction of strain-specific B-cell responses following the prime and boost immunisations.
Results Whereas priming immunisation induced the expected increase in strain-specific antibody titres, a second booster dose was strikingly ineffective at further increasing antibody titres. Similarly, priming immunisation induced strain-specific B-cells, but a second booster dose did not further enhance the B-cell response. Poor antibody responses were associated with male gender and cumulative cigarette exposure.
Conclusions Prime-boost, double-dose immunisation does not further improve influenza vaccine immunogenicity in previously vaccinated COPD patients. These findings underscore the need to design more effective vaccine strategies for COPD patients for influenza.
Abstract
Prime-boost, double-dose influenza immunisation does not further improve vaccine response in COPD patients, underscoring the need to design more effective vaccine strategies to overcome immune hyporesponsiveness https://bit.ly/3pd2Zas
Introduction
Prime-boost, double-dose vaccination is generally considered an effective strategy to provide protective immunity in susceptible populations, particularly the elderly and those with predisposing comorbidities such as chronic lung disease, where vaccines are less effective [1]. However, it remains widely underappreciated that COPD patients are more susceptible to respiratory viruses per se, and have intrinsically weaker immune responses to vaccinations against influenza and other pathogens [2] than their peers; accelerated “immuno-ageing/senescence” may contribute to this phenomenon [3]. This may particularly predispose COPD patients to acute influenza infections, subsequent bacterial pneumonia and life-threatening thromboembolic events in the year after recovery [4, 5].
In the elderly the immunosenescence that attenuates immunity to influenza vaccination can be overcome, in part, by higher dose and adjuvanted vaccines [6, 7]. Similarly, homologous prime-boost vaccination protocols, where second doses of the same vaccine are separated by 3–4 weeks, can strengthen humoral immune responses [1, 8, 9]. Much less is understood about vaccine responses in COPD [10, 11]. Antibody titre is conventionally used as a surrogate end-point in inactivated influenza vaccine trials [12], particularly because humoral rather than cellular immunity is mostly elicited by this technology [13, 14]. Of these antibodies, those against haemagglutinin (HA) are most often used, by convention, to measure response, although those against neuraminidase can also confer protection [15, 16]. We have previously reported that humoral immunity and seroconversion rates were markedly attenuated in COPD patients to the 2010 trivalent inactivated influenza vaccine against the pandemic (H1N1) 2009 strain (A/California/7/2009), a H3N2 strain (A/Perth/16/2009) and a B strain (B/Brisbane/60/2008) [11]. Such inactivated vaccines effectively elicit humoral antibody responses but typically fail to elicit effective CD8+ T-cell immunity. A recent Cochrane Review identified only one double-blinded randomised controlled trial on doubling vaccine dose in COPD [17]. In that study, which was performed in a predominantly vaccine-naïve population, a trivalent vaccine used at double the standard dose was found to be 76% effective in preventing influenza infections [18]. However, the effectiveness of higher dose, prime-boost immunisations in vaccine-experienced COPD patients receiving annual recommended vaccinations has not been reported.
In 2018 the Australian COPD Guidelines [19] were changed to recommend doubling influenza vaccination dose in COPD patients. This change in guidelines enabled us to study the effectiveness of doubling the standard dose of influenza vaccine using a prime-boost immunisation schedule in COPD patients in Melbourne and Brisbane who were already screened and enrolled in ongoing Australian National Health and Medical Research Council (NHMRC)-funded cohort studies on vaccine immunity. We assessed antibody titres and strain-specific B-cell responses. In contrast to previous reports we observed a striking lack of effectiveness of this immunisation protocol to further increase antibody titres in the majority of COPD patients. Our study suggests that the prime-boost strategy is unlikely to be effective, mitigating against undertaking larger randomised studies on this strategy, and underscores the need to specifically study vaccine effectiveness in COPD in order to develop more effective strategies.
Methods
Study population
Participants were recruited across two sites as a substudy of ongoing cohort studies on vaccine immunity at the Princess Alexandra Hospital (Brisbane, Australia) and The Royal Melbourne Hospital (Parkville, Australia) from the Melbourne Longitudinal COPD Cohort. Eligible participants were >55 years of age with a current clinical diagnosis of mild-to-very-severe COPD, and post-bronchodilator forced expiratory volume in 1 s (FEV1) <80% predicted and FEV1/forced vital capacity (FVC) ratio <0.7, with no COPD exacerbations in the 28 days prior to the study. Subjects were enrolled if: comorbidities (e.g. cardiac diseases (including ischaemic heart disease, cardiac arrhythmias and cardiac failure), diabetes mellitus and hypertension) were stable and well controlled, and use of inhaled or low-dose oral corticosteroid was stable in the 28 days preceding and remained stable post-vaccination. Exclusions were: invasive malignancy within the past 2 years, renal impairment (estimated glomerular filtration rate <40 mL·min−1·1.73 m−2), acute febrile illness with fever >38.5°C, hypersensitivity to egg proteins and use of oral prednisolone or equivalent ≥10 mg·day−1, or use of other immunosuppressive therapy. Patient characteristics are shown in table 1. Patients were also asked if they ever had doctor-diagnosed asthma and a proportion of patients were likely, therefore, to have COPD–asthma overlap, which is very common in the general community.
Demographics and baseline characteristics of the study population
Study design
Recruitment occurred preceding the southern hemisphere influenza season and written informed consent was obtained from each subject prior to commencement. Detailed clinical assessment, blood collection and spirometry were performed at day 0, prior to intramuscular administration of a single dose of the standard 2018 inactivated, quadrivalent, split-virion influenza vaccine (FluQuadri; Sanofi Pasteur, Paris, France) (immunisation protocol and sample collection shown in supplementary figure S1). The 2018 vaccine consisted of 15 μg HA each of: A/Michigan/45/2015 (H1N1) pdm09-like virus, A/Singapore/INFIMH-16-0019/2016 (H3N2)-like virus, B/Phuket/3073/2013-like virus and B/Brisbane/60/2008-like virus. Participants returned at day 28 post-vaccine for a second visit, where blood samples were taken prior to administration of a second vaccine dose. The first patient data were collected on 9 April 2018 and the last sample was collected on 29 August 2018.
Participants attended two further clinic visits on days 56 and 84 to provide blood samples. Blood samples were processed on the day of collection; peripheral blood mononuclear cells (PBMCs) and sera were stored at −80°C.
Annual vaccination against influenza, which is subsidised and widely available, is recommended for all COPD patients in Australia but not mandated. As a matter of record, the quadrivalent vaccines available prior to our study in 2016 comprised A/California/7/2009 (H1N1)-like virus, A/Hong Kong/4801/2014 (H3N2)-like virus (H3N2 is generally the most severe form of influenza A), B/Brisbane/60/2008-like virus and B/Phuket/3073/2013-like virus, and in 2017 comprised A/California/7/2009 (H1N1)-like virus, B/Brisbane/60/2008-like virus, B/Phuket/3073/2013-like virus and A/Michigan/45/2015 (H1N1)-like virus.
This study was conducted in accordance with the Declaration of Helsinki principles and the NHMRC Code of Practice, and is registered at the Australian New Zealand Clinical Trials Registry with identifier number ACTRN12620000954921. Ethical approval was granted by the local ethics committee for each site: The University of Queensland Human Ethics Research Office (clearance number: 2011000502), Metro South Health Human Research Committee (HREC/09/QPAH/297) and Royal Melbourne Hospital/Melbourne Health (HREC MH 2019:086).
Immunogenicity
Haemagglutination inhibition assay (HIA) was performed on receptor-destroying enzyme-treated sera, against components of each vaccine strain using microtitre techniques at the World Health Organization Collaborating Centre for Reference and Research on Influenza (Victorian Infectious Disease Reference Laboratory, Melbourne, Australia) [20]. Briefly, serial two-fold dilutions in PBS (1:10 to 1:1280) were incubated with 4 haemagglutinating units of the vaccine strain-specific influenza antigen and 1% turkey erythrocytes (H1N1 and B strains) or 1% guinea pig erythrocytes in the presence of oseltamivir (H3N2). Strain-specific HA antibody titres were calculated as the reciprocal of the highest dilution of sera that inhibited haemagglutination. Titres below the limits of detection (<10 or <20) were arbitrarily designated a value half the threshold of detection. The primary study end-point was seroconversion (defined as a ≥4-fold increase in haemagglutination inhibition antibody titre). Seroprotection (defined as a haemagglutination inhibition antibody titre ≥1:40) was a secondary end-point.
Strain-specific B-cell response
PBMCs were isolated using density gradient centrifugation and stored at −80°C until required. To determine the quantity and phenotype of B-cells elicited, influenza-specific B-cells were identified using recombinant HA probes and assessed by flow cytometry as described elsewhere [20]. Probes were developed for two 2018 southern hemisphere influenza vaccine strains: A/Michigan/45/2015 (H1N1) pdm09-like virus and B/Phuket/3073/2013-like virus. Due to the lack of availability of probes for the A/Singapore/INFIMH-16-0019/2016 (H3N2)-like virus and B/Brisbane/60/2008-like virus strains, B-cell data for these strains were not assessed. To phenotype strain-specific memory B-cells, CD19+IgD− B-cells were identified with recombinant HA probes, and CD27 and CD21 surface marker expression determined by flow cytometry to differentiate between classic memory B-cells (CD27+CD21+), activated memory B-cells (CD27+CD21−), naïve B-cells (CD27−CD21+) and atypical memory B-cell (CD27−CD21−, also referred to as “anergic” or “exhausted”) populations. The gating strategy and antibodies used are described in supplementary figure S3 and supplementary table S1, respectively. We have previously observed in our COPD cohort that inactivated vaccines do not elicit CD8 T-cell responses, so these were not measured in this study.
Statistical analysis
41 participants were recruited. Seven participants did not supply blood samples for all 4 days and one did not provide full demographic information, and were excluded from statistical analysis. For the remaining 33 study participants descriptive statistics were calculated separately for each vaccine strain. Antibody HIA titres were summarised as geometric mean titre with 95% confidence intervals. Antibody titres were log transformed and significant differences for each time-point compared with baseline determined by repeated measures ANOVA. Changes in mean percentages and phenotype of strain-specific B-cells were assessed with Wilcoxon's matched-pairs signed-rank test. Correlations between day 28 HIA titre and clinical characteristics, and day 28 HIA titre and strain-specific B-cells and titre, were assessed using Spearman's correlation coefficient (rs) for non-Gaussian distributions. With ability to seroconvert as the dependent variable, generalised linear models (GLMs) were used to identify factors independently associated with antibody response and any interactions between such factors. Descriptive statistical analyses were calculated with Prism version 8.4.2 (464) (GraphPad, San Diego, CA, USA). GLMs and clinical correlations were calculated using R version 3.5.2 (The R Foundation for Statistical Computing, Vienna, Austria). A p-value of <0.05 was considered to indicate statistical significance.
Results
Study participants
Characteristics of the 33 study participants are described in table 1. Most were aged in their 60s and 70s, and had moderate-to-severe COPD (mean FEV1 52.8% (95% CI 45.8–59.8%) predicted). 66% of participants were male and just over 30% were current smokers. Most participants had been vaccinated within the preceding 2 years; only four participants were influenza vaccine-naïve (table 1).
A second vaccination dose at day 28 does not improve strain-specific antibody titres
Vaccination with the 2018 seasonal quadrivalent influenza vaccine (15 μg HA per strain) resulted in the predictable increase in mean serum titres of A/H1N1-specific antibodies at day 28 (figure 1), although seroconversion rates were <40% across all vaccine strains (supplementary table S2). However, a second vaccination with the same dose at day 28 did not induce any additional increase in mean antibody titre at day 56 or day 84 (figure 1) nor did it improve seroconversion rates (supplementary table S2). In fact, mean antibody titre was lower at both these time-points than at day 28, demonstrating an inability of double-dose (sequential prime-boost) vaccination to maintain or augment antibody titres (figure 1 and supplementary figure S2). Similar response patterns were observed with the other vaccine strains (A/H3N2, B/Phuket and B/Brisbane): the second vaccine dose at day 28 did not induce any additional increase in mean antibody titres at day 56 or day 84 and did not prevent a decline in mean antibody titres at day 84 (figure 1). GLM analysis indicated that females were more likely to seroconvert than males, although this was only significant for the A/H1N1 strain (p=0.0278) (table 2). Stepwise GLM analysis using current smoking status, pack-years and FEV1/FVC ratio indicated that pack-years had more influence on ability to seroconvert than current smoking status or FEV1/FVC ratio. The negative association between pack-years and seroconversion was statistically significant for the A/H1N1 strain in univariate analysis (p=0.049); however, it was not significant when adjusted for current smoking status and FEV1/FVC ratio (table 2). Further details on seroconversion rates for each strain are provided in supplementary table S2.
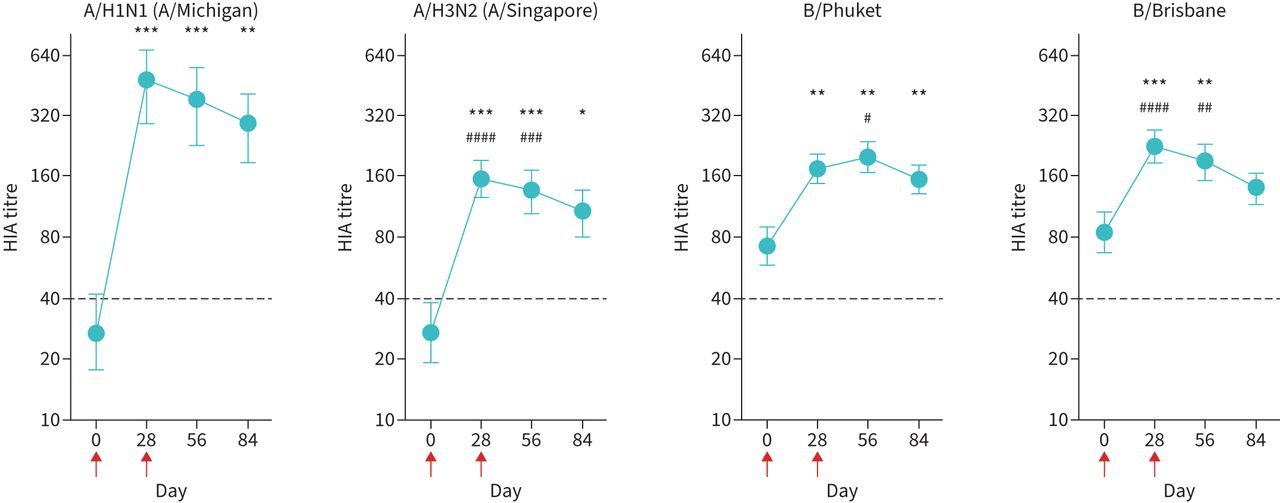
A second vaccination dose does not increase or sustain strain-specific antibody titres. Serum antibody titres were determined by haemagglutination inhibition assay (HIA). Red arrows indicate days of vaccinations. Data are presented as geometric mean titre with 95% confidence interval. Dotted horizontal black line indicates seroprotection titre (1:40). n=33. Statistical significance was determined by repeated interval ANOVA. *: p<0.05; **: p<0.01; ***: p<0.001 relative to day 0. #: p<0.05; ##: p<0.01; ###: p<0.001; ####: p<0.0001 relative to day 84.
Binomial generalised linear models (GLMs) determining factors influencing antibody response (seroconversion) for each vaccine strain
A second vaccination dose at day 28 does not improve the induction of strain-specific B-cells and does not alter the phenotype of circulating B-cells
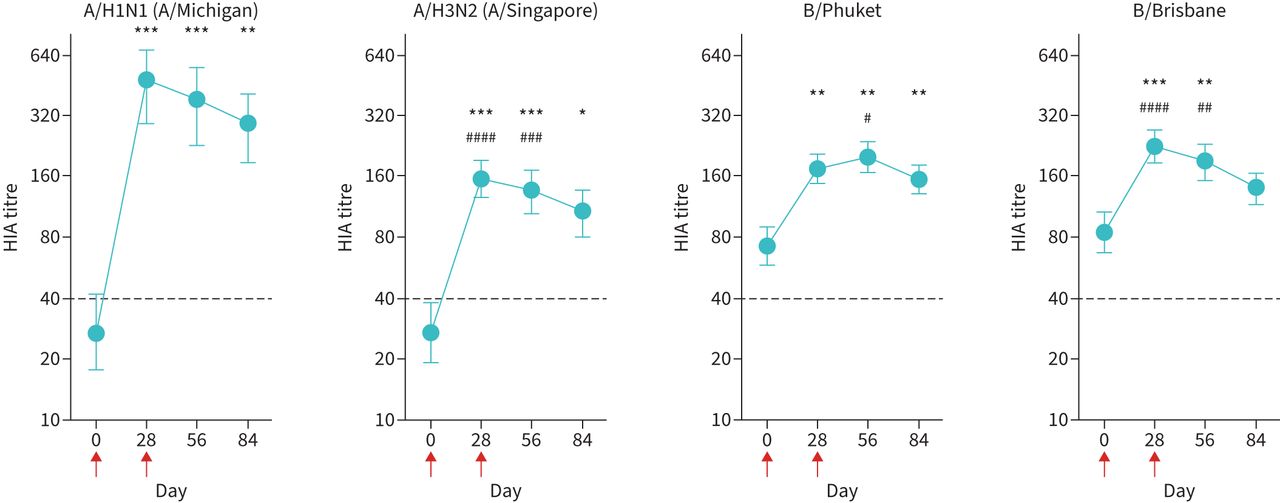
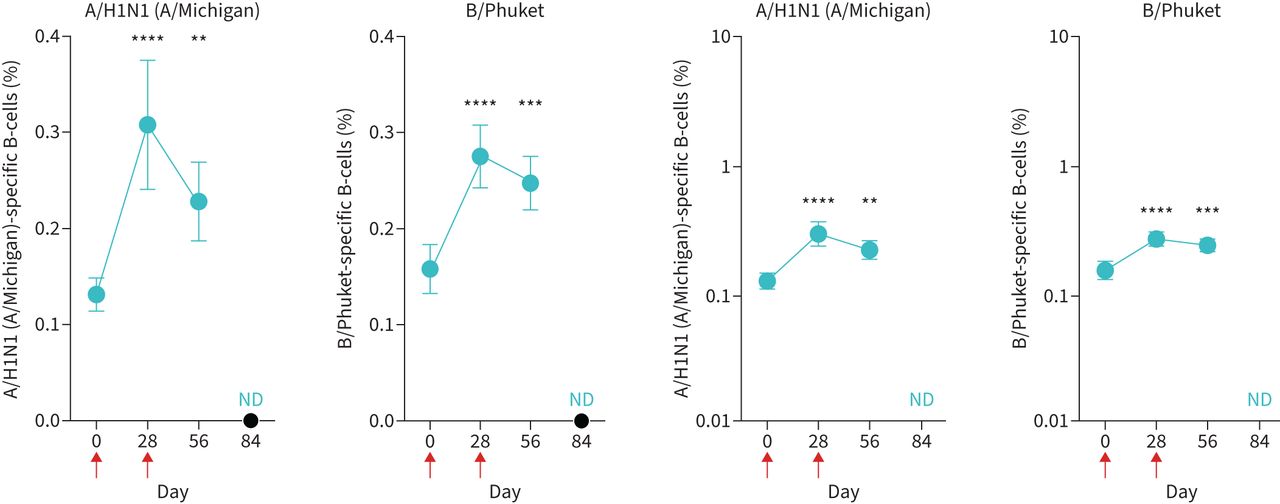
We next asked whether a second sequential vaccination altered induction of vaccine strain-specific B-cells. This involved the use of validated probes recognising type A (A/H1N1) and type B (B/Phuket) antigens [21–23]. Vaccination led to an increase in both A/H1N1- and B/Phuket-specific B-cells at day 28 (figure 2). However, a second vaccination at day 28 did not induce any additional B/Phuket-specific B-cells and was unable to maintain the A/H1N1-specific B-cells induced by the initial vaccination (figure 2). There were strong associations between specific antibody titres, and the proportions of A/H1N1-specific B-cells and B/Phuket-specific B-cells are shown in supplementary figure S4.

A second vaccination dose does not improve the induction of strain-specific B-cells. Peripheral blood mononuclear cells were stained with A/H1N1 (A/Michigan)- and B/Phuket-specific probes to determine the percentage of cells within the blood CD19+IgD− B-cell population by FACS. Red arrows indicate days of vaccinations. Data are presented as mean±sem. n=33. Note difference in y-axis scales to accomodate data: left panels, linear scale; right panels, log scale. ND: not determined. Statistical significance was determined using Wilcoxon's matched-pairs signed-rank test. **: p<0.01; ***: p<0.001; ****: p<0.0001 relative to day 0.
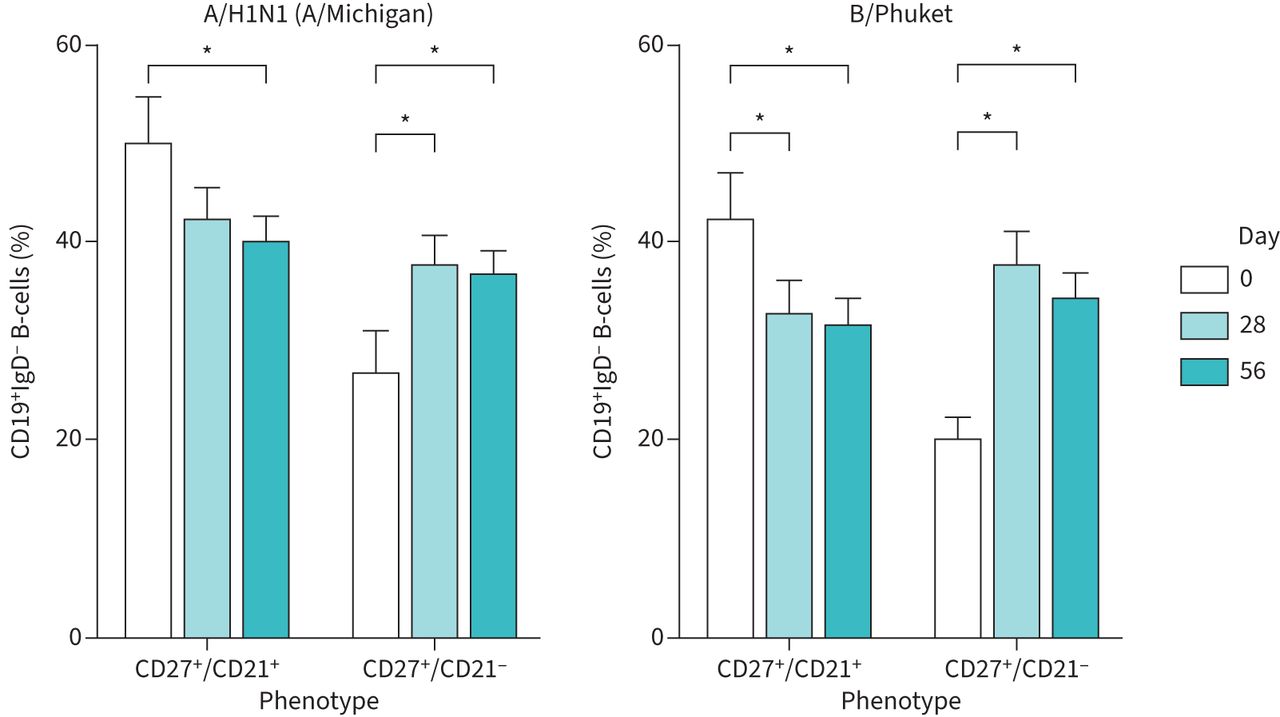
Similarly, while initial vaccination at day 0 induced an increase in activated memory B-cells and a concomitant reduction in the percentage of A/H1N1-specific classic memory B-cells at day 28, a second vaccination at day 28 did not induce any additional effects on the percentage of either population at day 56 (figure 3). Vaccination had minimal effects on the relatively low percentages of naïve and atypical memory B-cell populations (supplementary figure S5).

{kind=link}
{kind=link}
{kind=link}
A second vaccination dose does not affect vaccine-induced changes in B-cell memory. CD19+IgD− peripheral blood mononuclear cells were stained with A/H1N1 (A/Michigan)- and B/Phuket-specific probes, and cells differentiated on the basis of CD27 and CD21 expression by FACS. n=33. Statistical significance was determined using Wilcoxon's matched-pairs signed-rank test. *: p<0.05.
Discussion
In the present study we evaluated the outcome of delivering double the usual dose of seasonal influenza vaccine to COPD patients using a prime-boost immunisation schedule to determine whether this might enhance immunogenicity. Priming immunisation induced a significant increase in strain-specific antibody titres across all four vaccine strains. However, our results reveal a striking lack of effectiveness of the second “boost” arm of this strategy, which was unable to further increase antibody titres or B-cell responses. These results contrast markedly with a previous report in influenza vaccine-naïve COPD patients; however, that study did not use a prime-boost vaccination schedule [18].
Notwithstanding the failure of the boost to enhance immunity it is important to note that we, as have others, did observe a positive humoral response to the priming vaccination in many patients. Given the susceptibility of COPD patients to influenza, post-infection pneumonia and its sequalae, our results clearly support the continued use of influenza vaccination in this population. Our study was not designed to understand if a prime-boost, double-dose strategy actually prevents influenza infection, as such studies require very large cohorts of thousands of patients over an extended period of time.
In our study we observed that the majority of patients had some increase in antibody titre and up to 36% reached the 4-fold increase conventionally regarded as seroconversion. Of note, several of the antigens present in the 2018 vaccine were also present the 2016 and 2017 vaccine (as detailed earlier) and this may have influenced the 2018 response to those antigens, e.g. by yielding a higher baseline and therefore an apparently smaller change in titre. These antibody responses were mirrored by increases in vaccine strain-specific B-cells. Immunologically, it is known that inactivated influenza vaccines activate both B-cells and T follicular helper (TFH) cells to induce antibody-secreting cells and increases in serum antibody titres [21]. The Koutsakos et al. [21] study in a general population of healthy adults demonstrated that inactivated influenza vaccinations induce three main B-cell responses comprising a transient CXCR5−CXCR3+ antibody-secreting B-cell population, CD21hiCD27+ memory B-cells and CD21loCD27+ B-cells. Activation of circulating TFH cells was shown to correlate with the development of both CD21lo and CD21hi memory B-cells [21]. In our study, while the trend was not marked or statistically significant, many patients actually showed a small decrease in antibody titre after boost. Previous studies have observed a negative correlation between prior antibody levels and subsequent B-cell responses. Whether repeat immunisation to the same epitope leads to clinically important decreases in humoral immunity, and if so what mechanism(s) might cause this, are areas of current controversy.
In order to better understand correlates of poor humoral responses, we analysed multiple potential determinates using a GLM statistical approach. Males seroconverted less readily relative to females. Interestingly, current smoking was not independently associated with seroconversion, although overall pack-year history was negatively correlated. Active smoking, or even exposure to passive cigarette smoke, has repeatedly been shown to enhance susceptivity to influenza and other viruses, and this is thought to reflect mucosal immune suppression. It is, however, likely that long exposure to smoke reflected in pack-year history damages the immune system to an extent that active smoking no longer appears as a risk factor [24]. Age, a well-established determinant of weak vaccine responses, was not significantly associated with seroconversion, most likely because all of our patients were elderly and in a narrow age range. It is known that TFH cell responses are required to consolidate high-affinity antibody production and that these wane with age [25], although it is not known if COPD specifically weakens this compartment.
Our study has several limitations. Key among these are that our study, for ethical reasons, was constrained to an open-label design, and that it is not possible to measure actual protection against influenza infection for studies of this size and duration. Protection studies require very large cohorts of thousands of subjects and a lengthy duration. We did not measure antibodies at the respiratory mucosal surfaces nor could we assess the anatomical distribution of memory cells which, in post-mortem studies, have shown a predilection to redistribute from blood to lung tissue. While it is conceivable that COPD patients might show favourable changes in antibody titres in these compartments, but not in the circulation, this seems unlikely. Even in healthy people inactivated influenza vaccines produce relative weak and usually transient increases in mucosal antibodies [26].
All of the patients in our study met Global Initiative for Chronic Obstructive Lung Disease (GOLD) criteria for the diagnosis of COPD; however, a proportion of patients reported a past diagnosis of asthma, which has a high population prevalence in Australia, and some were likely to have at least some features of asthma–COPD overlap, common in “real-world” cohorts. Our study was not sufficiently powered to determine if responses to double-dose influenza vaccine differed in patients with pure COPD versus those with both COPD and asthma. This is a limitation of our study and warrants further investigation in larger studies. However, it is noteworthy that our recent much larger study of single-dose influenza vaccination found that neither COPD nor asthma were associated with seroconversion and seroprotection [27].
We would also like to explicitly state that our results are not generalisable to adenoviral or RNA vaccine technologies being employed against severe acute respiratory syndrome coronavirus 2. For example, inactivated influenza vaccines produce no, or negligible, anti-viral T-cell responses.
Despite these limitations our study clearly demonstrates that a prime-boost, double-dose immunisation schedule is not effective in increasing humoral immunity in COPD. Given that COPD patients are a large and highly vulnerable population, our studies highlight the need to specifically study this population and, more importantly, to develop much more effective influenza vaccination strategies in this group.
Supplementary material
Supplementary Material
Please note: supplementary material is not edited by the Editorial Office, and is uploaded as it has been supplied by the author.
Supplementary material 00641-2021.SUPPLEMENT
Footnotes
Provenance: Submitted article, peer reviewed.
This study is registered at the Australian New Zealand Clinical Trials Registry with identifier number ACTRN12620000954921. Individual participant data that underlie the results reported in this article, after de-identification (text, tables, figures and appendices), and the study protocol, statistical analysis plan and analytic code will be available beginning 3 months and ending 5 years following article publication, to researchers who provide a methodologically sound proposal, to achieve aims in the approved proposal. Proposals should be directed to Gary Anderson (gpa{at}unimelb.edu.au). To gain access, data requesters will need to sign a data access agreement.
Author contributions: All authors met the International Committee of Medical Journal Editors criteria for authorship. Specifically they all: made substantial contributions to the conception or design of the work; or the acquisition, analysis or interpretation of data for the work; and drafted the work or revising it critically for important intellectual content; and gave their final approval of the version to be published; and agree to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Conflict of interest: All authors declare no conflict of interest.
Support statement: The study was funded by the National Health and Medical Research Council of Australia (grant number APP1081433). Funding information for this article has been deposited with the Crossref Funder Registry.
- Received November 16, 2021.
- Accepted August 1, 2022.
- Copyright ©The authors 2023
This version is distributed under the terms of the Creative Commons Attribution Non-Commercial Licence 4.0. For commercial reproduction rights and permissions contact permissions{at}ersnet.org
References










