





Abstract
Background Electronic cigarettes (ECs) are electronic aerosol delivery systems composed of nicotine and various chemicals, which are widely used to facilitate smoking cessation. Although ECs are considered safer than cigarettes, they do, however, contain chemical toxicants, some of which may interact with cells of the host's innate immune system of which neutrophils constitute a key component.
Methods The current study was designed to compare the effects of aqueous EC aerosol extracts (ECEs; with or without nicotine) with those of cigarette smoke extract (CSE) on neutrophil and platelet reactivity in vitro. Neutrophil reactivity is characterised by the generation of reactive oxygen species (ROS), degranulation (elastase release) and the release of extracellular DNA (neutrophil extracellular trap (NET) formation: NETosis), which were measured using chemiluminescence, spectrophotometric and microscopic procedures, respectively. Platelet reactivity was measured according to the magnitude of upregulated expression of the adhesion molecule CD62P on activated cells using a flow cytometric procedure.
Results Exposure of neutrophils to either ECEs or CSE caused a significant inhibition of ROS generation and elastase release by N-formyl-l-methionyl-l-leucyl-l-phenylalanine (1 µM)-activated neutrophils. Pre-treatment of neutrophils with CSE also resulted in a marked attenuation of phorbol 12-myristate 13-acetate (6.25 nM)-mediated release of extracellular DNA, which was unaffected by the ECEs. Similarly, CSE, but not the ECEs, inhibited the expression of CD62P by platelets activated with ADP (100 µM).
Conclusions These observations suggest that ECE aerosols may inhibit some of the immuno-protective activities of neutrophils such as ROS production and elastase release by activated cells, the effect of which was not enhanced by inclusion of nicotine. The inhibitory effects of CSE were significantly more pronounced than those of ECEs, especially so for suppression of NET formation and platelet activation. If operative in vivo, these harmful immunosuppressive effects of ECEs may compromise intrinsic pulmonary antimicrobial defence mechanisms, albeit less so than cigarette smoke.
Abstract
Electronic cigarette (EC) vapours, similarly to cigarette smoke, adversely affect the innate immune system, which could potentially predispose EC users to respiratory infections https://bit.ly/3M2KIJl
Introduction
Electronic cigarettes (ECs) are electronic nicotine delivery systems that allow users to inhale an aerosol, which typically contains nicotine, flavourants and other additives [1]. Originally entering the market in 2003, these devices were patented internationally more than a decade ago, and since then have gained popularity among both smokers and non-smokers of all ages, particularly adolescents [2]. EC use may be considered as a useful smoking cessation strategy, although evidence for this has been regarded as “unconvincing” by some [3, 4] and of “moderate certainty” by others [5]. Although EC use is particularly popular among non-smoking adolescents, concerns exist that it may serve as a “gateway drug” in this group [3, 4]. Various chemicals and potential toxicants have been identified in EC aerosols and although the concentrations of these have been found to be lower than in cigarette smoke, they may not be completely risk free [6, 7].
Neutrophils are phagocytic leukocytes that constitute the first line of the host cellular innate immune response against invading pathogens [8, 9]. They capture and destroy microorganisms via a number of mechanisms, including phagocytosis, generation of reactive oxygen species (ROS), degranulation and neutrophil extracellular trap (NET) formation (NETosis) [8, 9]. Defects in these functions can compromise host defences, leading to overwhelming infections with bacteria and fungi, underscoring their importance to respiratory defences [10].
Platelets play a central role in haemostasis; however, they have also been found to exert a range of immune-related functions, including interactions with neutrophils, which amplify the pro-inflammatory activities and transendothelial migration of these cells [11]. Platelet–neutrophil complex formation is mediated by P-selectin (CD62P) and its ligand P-selectin glycoprotein ligand-1 (PSGL-1), which enable first contact and initiate intracellular signalling [12]. These complexes are present in a wide range of inflammatory conditions and infections [13].
Cigarette smoke condensate (CSC) has previously been shown to attenuate NET formation and oxygen consumption (as an indirect measure of ROS production) by phorbol 12-myristate 13-acetate (PMA)-activated human neutrophils in vitro [14]. Although consistent with a suppressive effect of CSC on the protective activities of human neutrophils, the question arose as to whether exposure to EC aerosol would also affect these cells. In the current study, the effects of aqueous EC aerosol extracts (ECEs) on neutrophil functions and certain indices of platelet activation were evaluated in vitro. Cigarette smoke extract (CSE) was used as a control.
Methods
Study details and participants
This was a prospective, experimental laboratory-based study designed to investigate and compare the effects of CSE and ECEs on the functions of isolated human blood neutrophils and platelets in vitro.
Permission to undertake this study and draw blood from healthy, adult human volunteers was granted by the Research Ethics Committee of the Faculty of Health Sciences, University of Pretoria (Pretoria, South Africa) in full compliance with the World Medical Association Declaration of Helsinki 2013 (approval: 123/2018).
Participants included in the study were healthy adults (n=19; mean±sd age 34±14 years; male:female ratio 5:14). Exclusion criteria included: 1) those with chronic medical conditions and anaemia; 2) current and ex-smokers; and 3) any potential donor deemed unwell by the qualified nursing sister in attendance on the day of venepuncture. Informed consent was obtained from all participants in the study prior to the blood draw.
Chemicals and reagents
Three different extracts were used in this study: aqueous extracts derived from the aerosols of 1) nicotine-free ECs (ECE−Nico); 2) nicotine-containing ECs (ECE+Nico); and as a reference product, 3) aqueous extracts derived from cigarette smoke (CSE). The method used to obtain these extracts is described in the following sections. Unless indicated, all other chemicals and reagents were obtained from Sigma (St Louis, MO, USA).
Preparation of ECE and CSE
EC vapour was generated using the Twisp Clearo Pod (Twisp, Cape Town, South Africa; subsequently acquired by British American Tobacco) filled with tobacco-flavoured e-liquid without and with nicotine (18 mg·mL−1). The pod was attached to an apparatus which pumped the EC vapour into collection tubes containing 10 mL Hanks’ balanced salt solution (HBSS; indicator-free, pH 7.4). The pump delivered a 5-s puff of EC vapour followed by a 5-s resting interval. Each cycle spanned a 5-min period delivering 30 puffs per 10 mL sample.
CSE was prepared from the combustion of commercially available medium tar (10 mg tar and 0.8 mg nicotine) Marlboro Red (Phillip Morris, Richmond, VA, USA) cigarettes to generate tobacco smoke that was passed through HBSS with formation of a condensate that was diluted to 10% and frozen at −80°C for use later. The nicotine concentration of the condensate was ∼9 µg·mL−1. The aforementioned protocols were adapted from those described by van Zyl-Smit et al. [15] and Chang et al. [16].
Preparation of neutrophils
Neutrophils were isolated from heparinised venous blood (5 U preservative-free heparin·mL−1 blood) from healthy adult volunteers. Neutrophils were separated from mononuclear leukocytes by centrifugation on Histopaque-1077 cushions at 400×g for 25 min at room temperature. The resultant pellets were suspended in PBS (0.15 M, pH 7.4) and sedimented with 3% gelatine to remove most of the erythrocytes. Following centrifugation (280×g at 10°C for 10 min), residual erythrocytes were removed by selective lysis with 0.83% ammonium chloride at 4°C for 10 min. The neutrophils, which were routinely of high purity (>90%) and viability (>95%), determined by flow cytometric procedures, were resuspended to 107 mL−1 in PBS and held on ice until used.
Measurement of ROS
Generation of ROS was measured using lucigenin (bis-N-methyl-acridinium nitrate)- and luminol (5-amino-2,3-dihydro-1,4-phthalazine dione)-enhanced chemiluminescence procedures that predominantly detect superoxide and ROS produced by the myeloperoxidase/H2O2/halide system, respectively [17]. Briefly, neutrophils (106 cells) were pre-incubated for 10 min at 37°C without or with the extracts (50%) in 900 µL HBSS containing either lucigenin (0.2 mM) or luminol (0.1 mM), followed by addition of either 100 μL HBSS (unstimulated control systems) or the chemoattractant N-formyl-l-methionyl-l-leucyl-l-phenylalanine (FMLP; 1 μM). Chemiluminescence was measured using a Lumac Biocounter (Model 2010; Lumac Systems, Titusville, FL, USA). The final volume in each vial was 1 mL and the results, expressed as relative light units (RLU), represent the peak values for FMLP-activated systems that were reached 40–50 s after addition of the stimulant.
Elastase release
Neutrophil degranulation was measured by quantifying the release of the primary granule enzyme elastase. Neutrophils were incubated at a concentration of 2×106 mL−1 in HBSS without and with the various extracts (50%) for 10 min at 37°C. FMLP (1 µM) in combination with cytochalasin B (1 µM final) was added to the cells for 10 min at 37°C. The tubes were then transferred to an ice bath and the cells centrifuged at 400×g for 5 min. The supernatants were decanted and assayed for elastase using a micromodification of a standard colorimetric procedure [18]. Briefly, 125 µL supernatant was added to the elastase substrate N-succinyl-l-alanyl-l-alanyl-l-alanine-p-nitroanilide (3 mM in dimethyl sulfoxide (DMSO)) in 0.05 M Tris–HCl (pH 8.0) and elastase activity monitored spectrophotometrically at a wavelength of 405 nm.
Spectrophotometric detection of NETs
Following pre-incubation of neutrophils for 10 min at 37°C, the extracts (50%) or an equal volume of HBSS (2 mL, control) were added to the neutrophils (4×106 cells in 4 mL HBSS) and the cells then activated with phorbol 12-myristate 13-acetate (PMA; 6.25 ng·mL−1 final). Following 90 min incubation at 37°C, the cells were pelleted by centrifugation and 3 mL of the supernatant mixed with 2 µL of the DNA-binding fluorophore Sytox Orange (5 µM final; Life Technologies, Eugene, OR, USA) in reaction cuvettes. The cuvettes were then transferred to the cuvette holder of a Hitachi 650 10S fluorescence spectrophotometer (Hitachi, Tokyo, Japan) and fluorescence measured with the excitation and emission wavelengths set at 530 and 590 nm, respectively. Results are expressed as metered fluorescence units (MFU).
Microscopic detection of NETs
Neutrophil suspensions (0.6×105 cells in 250 µL) were allowed to adhere to glass coverslips for 30 min. Extracts (50%) or an equal volume of HBSS (control) were added to the adherent cells, which were then activated with PMA (6.25 ng·mL−1 final) before being incubated for 120 min at 37°C/5% CO2. The cells were then fixed using 4% paraformaldehyde for 25 min, washed three times in PBS, and blocked with HBSS containing 5% goat serum and 5% bovine serum albumin for 30 min at 37°C. Coverslips were subsequently incubated overnight with polyclonal rabbit anti-histone H4 antibody (Merck, Darmstadt, Germany). The next day, coverslips were washed three times in PBS and exposed to Alexa Fluor 488-conjugated goat anti-rabbit antibody (Life Technologies) for 60 min at 37°C and washed three times in PBS before being stained for the presence of DNA using 4′,6-diamidino-2-phenylindole (DAPI; Life Technologies) for 30 min. The coverslips were mounted on microscope slides using anti-fade mounting fluid and the NETs were visualised by confocal microscopy using a Zeiss LSM800 Meta confocal microscope (Zeiss, Oberkochen, Germany) with a ×20 objective and a pinhole set to 1 Airy. The appropriate excitation and emission wavelengths were used for Alexa Fluor 488 and DAPI. Representative photomicrographs were taken.
Neutrophil viability
Neutrophil viability was measured after a 10- or 90-min exposure at 37°C to the extracts (50%). Neutrophil (106 mL−1) viability was assessed using a flow cytometry procedure based on propidium iodide dye exclusion (50 μg·mL−1; DNA Prep-Stain; Beckman Coulter, Miami, FL, USA). Results are expressed as the percentage of viable cells of total cells.
Preparation of platelet-rich plasma and detection of platelet CD62P expression
To prepare platelet-rich plasma (PRP), blood (anti-coagulated with 5 U preservative-free heparin·mL−1 blood) was centrifuged at 250×g for 10 min at room temperature within 15 min of venepuncture and the erythrocyte- and leukocyte-free upper layer of PRP was decanted and platelet activation measured according to the magnitude of expression of CD62P.
Platelet activation was measured by flow cytometry as the proportion of CD42a+ platelets expressing the α-granule-derived adhesion molecule CD62P. For these experiments, PRP (20 µL) was added to 980 µL HBSS and incubated for 10 min at 37°C in the presence of the different extracts (50%) or an equal volume of HBSS as control (0.5 mL). Subsequently, ADP (100 μM final; platelet P2Y1/P2Y12 receptor agonist) or an equal volume of HBSS (background control for spontaneous platelet activation) was added. Platelets were incubated for 5 min at 37°C and processed immediately thereafter for analysis by flow cytometry. The platelets were stained with 5 µL each of a murine anti-human platelet CD42a–phycoerythrin (PE)-labelled monoclonal antibody (Becton Dickenson, San Jose, CA, USA) and an anti-human CD62P–fluorescein isothiocyanate-labelled monoclonal antibody (Beckman Coulter) to detect the total and activated platelet populations, respectively. After 15 min incubation in the dark, the samples were analysed on a Gallios flow cytometer (Beckman Coulter) and the results expressed as the percentage of activated platelets with 50 000 cells interrogated during each measurement [19].
Neutrophil–platelet aggregate formation
Neutrophil–platelet (NP)-enriched buffy coats were prepared from heparinised human blood by sedimentation at 37°C. For these experiments buffy coat (30 μL) was added to 970 μL HBSS and incubated for 10 min at 37°C in the presence of the different extracts (50%) or an equal volume of HBSS (0.5 mL, control). This was followed by the addition of ADP (100 μM) or an equal volume of HBSS (background) and incubation for a further 5 min at 37°C. Cells were processed immediately thereafter for flow cytometry. Cell suspensions were stained with 5 μL murine anti-human fluorochrome-labelled monoclonal antibodies directed against CD16–PE cyanin 5 (Beckman Coulter) to detect neutrophils, CD42a–PE (Becton Dickenson) to detect platelets and CD45–Krome Orange (Beckman Coulter) to detect total leukocytes for 15 min at room temperature. This was followed by analysis of the various cell suspensions at a slow flow rate using a Gallios flow cytometer (Beckman Coulter). NP interactions were determined according to the CD16+/CD42a+ coexpression profiles of CD45+ leukocytes and the results expressed as the relative mean fluorescence intensities of CD42a expression of these NP aggregates [20].
Expression of results and statistical analysis
The results of each series of experiments are expressed as the mean and standard deviation with the numbers of different donors clearly indicated. Statistical analyses were performed using InStat and Prism 5 (GraphPad, San Diego, CA, USA) with levels of statistical significance calculated using the Mann–Whitney U-test for comparison of unpaired, non-parametric data. A two-tail p-value of <0.05 was considered significant.
Results
Production of ROS
To assess the effect of the extracts on the generation of ROS by neutrophils after stimulation, lucigenin- and luminol-enhanced chemiluminescence responses were measured after pre-incubation with the extracts followed by FMLP-dependent activation (figure 1). Treatment of the cells with the different extracts resulted in significant inhibition of the FMLP-dependent generation of ROS, with the strongest inhibition evident in CSE-exposed neutrophils. The inhibitory responses observed with ECE in the absence and presence of nicotine (ECE−Nico and ECE+Nico, respectively) were statistically similar, suggesting that the magnitude of inhibition was not augmented by the presence of nicotine in ECE, Thus, CSE and ECE both inhibit ROS production by activated neutrophils. The mean percentage inhibition of lucigenin-enhanced chemiluminescence (and luminol-enhanced chemiluminescence in brackets) was 55% (87%), 46% (58%) and 36% (59%) for systems containing CSE, ECE−Nico and ECE+Nico, respectively (p<0.05).

Effects of cigarette smoke extract (CSE) and aqueous electronic cigarette aerosol extracts without nicotine (ECE−Nico) and with nicotine (ECE+Nico) on lucigenin- and luminol-enhanced chemiluminescence responses of neutrophils activated by N-formyl-l-methionyl-l-leucyl-l-phenylalanine (FMLP; 1 µM). Results are presented as mean±sd peak chemiluminescence in relative light units (RLU) measured 30–50 s after the addition of FMLP. For lucigenin-enhanced chemiluminescence (left graph, n=7), the absolute values for unstimulated neutrophils (background) and for cells activated with FMLP were 1780±1184 and 7063±3554 RLU, respectively. The corresponding values for luminol-enhanced chemiluminescence (right graph, n=6) were 4875±3413 and 106 001±35 541 RLU, respectively. *: p<0.05 and #: p<0.05 for comparison with FMLP-activated cells.
Elastase release
To assess the effects of the extracts on elastase release by activated neutrophils, isolated cells were pre-treated with CSE, ECE−Nico or ECE+Nico before being activated with FMLP/cytochalasin B. Elastase expression in the supernatant was quantified (table 1). The data show that the robust release of elastase in response to FMLP/cytochalasin B was significantly inhibited by CSE (45%), as well as both ECE treatments (28% ECE−Nico and 20% ECE+Nico). However, CSE treatment was most potent with respect to elastase release. As seen with ROS production, the absence of nicotine in the ECE treatments did not alter the inhibitory effect of ECE, suggesting that nicotine is not active in inhibiting neutrophil activation. Thus, elastase secretion was inhibited by ECE and CSE.
Effects of cigarette smoke extract (CSE) and aqueous electronic cigarette aerosol extract without nicotine (ECE−Nico) and with nicotine (ECE+Nico) on the release of elastase from neutrophils activated with N-formyl-l-methionyl-l-leucyl-l-phenylalanine (1 µM)/cytochalasin B (1 µM), as well as the effects of the extracts on phorbol 12-myristate 13-acetate (PMA; 6.25 mg·mL−1)-activated release of extracellular DNA
Spectrophotometric detection of NETs
To assess if the extracts affected NET production by neutrophils, the presence of extracellular DNA was quantified by spectrophotometry (table 1). PMA activation of neutrophils induced a significant increase in the amount of extracellular DNA, which was significantly inhibited by pre-treatment of cells with CSE (463 versus 21 MFU). Interestingly, pre-treatment with ECE−Nico or ECE+Nico did not inhibit NETosis as evidenced by the high levels of extracellular DNA (357 and 482 MFU). Thus, exposure to CSE, but not ECE, caused significant inhibition of NETosis by PMA-activated neutrophils.
Microscopic detection of NETs
To confirm the effects of CSE and ECE on the induction of extracellular DNA or NETosis after PMA stimulation of neutrophils, confocal microscopy was performed to visualise the extracellular DNA. Histone H4 staining detected significant NET formation in PMA-stimulated neutrophils (figure 2b) compared with non-stimulated neutrophils (figure 2a). Strikingly, cells treated first with CSE before PMA stimulation did not exhibit the extensive NET formation seen in control cells treated with PMA only, even though some histone H4 staining within the cells themselves was evident (figure 2c), which is similar to the staining observed in the untreated cells. Interestingly, ECE-exposed cells did form NETs, with ECE+Nico presenting less than ECE−Nico, similar to the data obtained in the NETosis spectrophotometry analysis. Overall, only CSE-treated cells showed an almost complete lack of NETs, while ECE-treated cells still presented NETs.

Confocal microscopy images of neutrophil extracellular trap formation. Neutrophils were stimulated with a) Hanks’ balanced salt solution alone, b) phorbol 12-myristate 13-acetate (PMA) alone, c) cigarette smoke extract followed by PMA, d) electronic cigarette aerosol extract (ECE) without nicotine (ECE−Nico) followed by PMA and e) ECE with nicotine (ECE+Nico) followed by PMA. Scale bar: 50 µm. Extracellular DNA is visualised using citrullinated histone H4 antibody and Alexa Fluor 488 (green), as well as 4′,6-diamidino-2-phenylindole for total DNA (blue).
Neutrophil viability
To assess whether the inhibition of neutrophil activation after exposure to CSE or ECE was due to a decrease in cell viability, cells were exposed to the extracts for 10 or 90 min and viability was assessed. Control cells had viabilities after 10 and 90 min of 97.5±1.7% and 97.5±1.0%, respectively, while exposure to CSE (97.5±1.8% and 97.3±0.7%, respectively), ECE−Nico (97.5±1.2% and 98±0.5%, respectively) or ECE+Nico (97.3±1.4% and 98.4±0.4%, respectively) did not affect neutrophil viability. Thus, the effects observed on neutrophil activation could not be ascribed to changes in viability.
Expression of CD62P by PRP
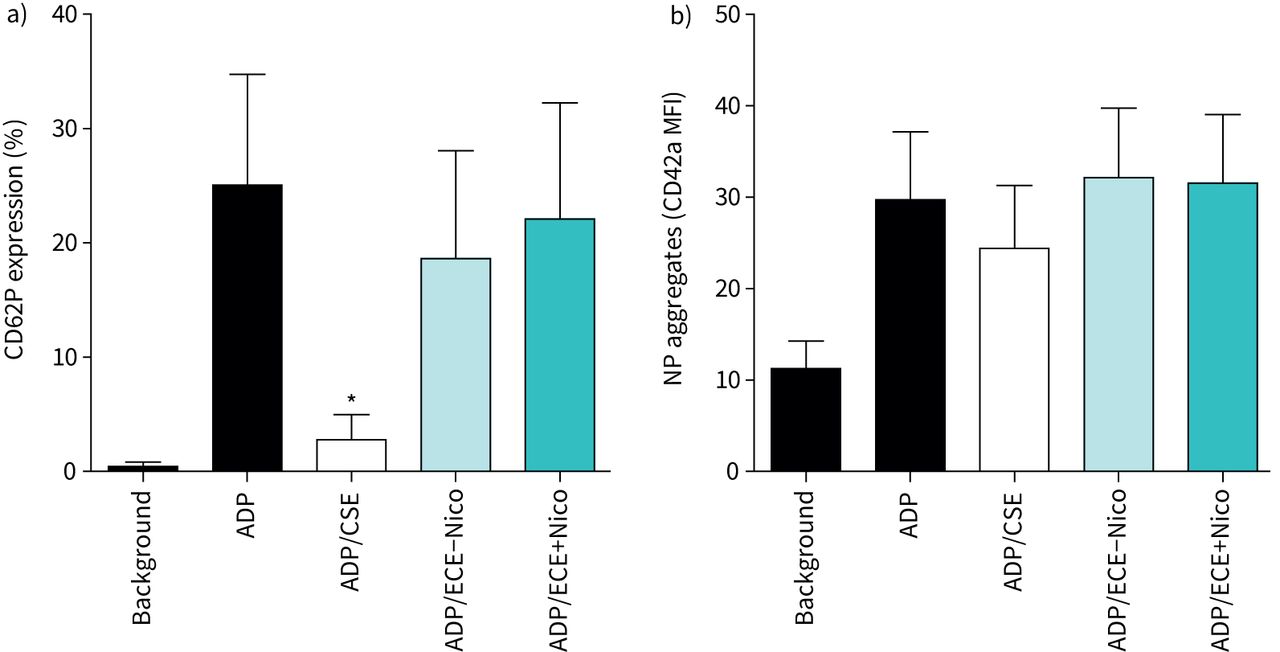
While we observed that neutrophil activation was negatively affected by CSE or ECE, we now investigated whether this also holds true for platelet activation. To assess platelet activation, expression of CD62P was measured following activation of the cells with ADP. Analysis of CD62P expression showed that stimulation with ADP resulted in a significant increase in expression of the adhesion molecule, which was significantly attenuated by exposure of the cells to CSE to a level comparable to that of non-activated platelets (figure 3a). In contrast, ECE treatment did not affect CD62P expression levels, demonstrating that only CSE inhibited platelet activation.
{kind=link}
{kind=link}
{kind=link}
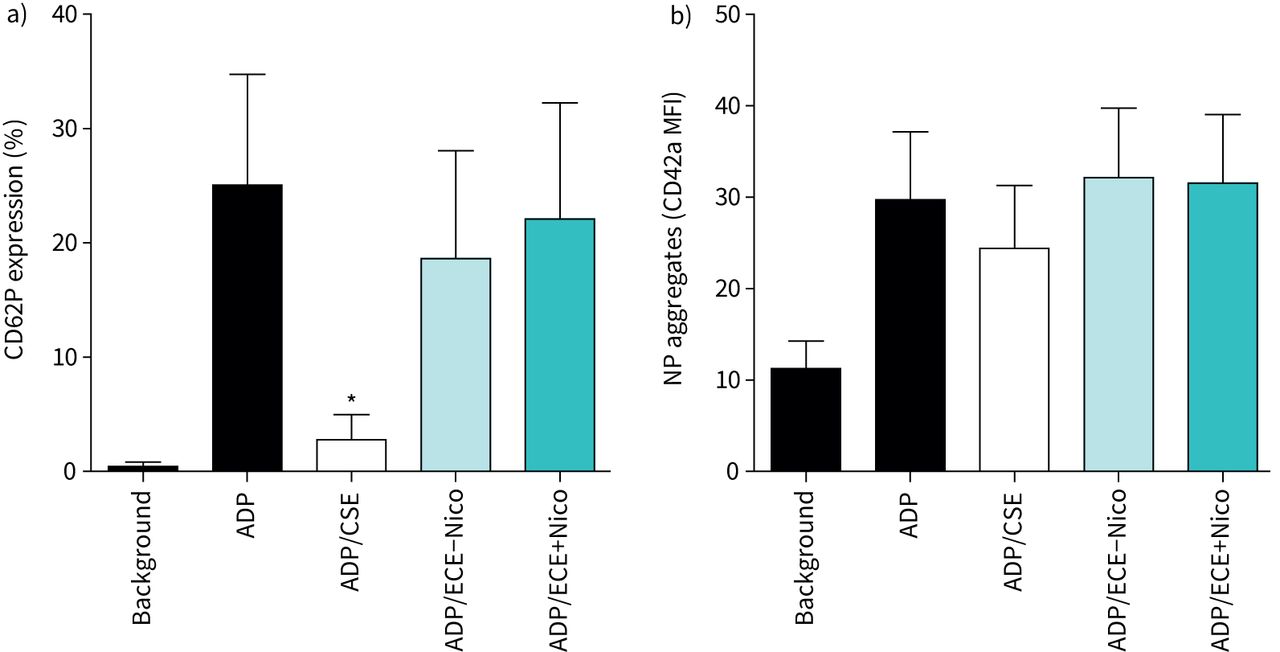
a) Effects of cigarette smoke extract (CSE) and electronic cigarette aerosol extracts without nicotine (ECE−Nico) and with nicotine (ECE+Nico) on the magnitude of platelet CD62P expression following activation with ADP (100 µM). Results are presented as mean±sd percentage CD62P expression (n=6). The absolute values for unstimulated neutrophils (background) and for cells activated with ADP in the absence of extracts were 0.44±0.39% and 25.0±9.7%, respectively. *: p<0.05 for comparison of CSE (50%) with the drug-free control system. b) Effects of CSE, ECE−Nico and ECE+Nico on ADP-activated heterotypic neutrophil–platelet (NP) aggregation. Results are presented as mean±sd CD42a mean fluorescence intensity (MFI) (n=5). The aggregates assessed were positive for coexpression of CD16, CD42a and CD45.
NP aggregate formation
NP aggregation was assessed by quantifying CD42a expression in the presence of CD16 expression. While stimulation of the cell mix with ADP led to a significant increase in aggregation, pre-treatment with either CSE or ECE did not alter this aggregation, indicating that aggregation was not affected by either extract (figure 3b).
Discussion
Upon heating, ECs generate aerosols which contain several toxicants potentially hazardous to respiratory health. These include the vapourising agents, glycerol and propylene glycol and their thermal degradation products (i.e. carbonyls), as well as heavy metals. These toxicants are common to most brands of ECs, while the type and concentrations of others, such as chemical flavourants and nicotine, vary according to the different brands of ECs [21]. However, compared with cigarette smoke emissions, aerosols of ECs containing recent design features show substantially lower levels of major cigarette smoke toxicants [22]. Nevertheless, given their potential to adversely affect respiratory health, additional insights are required to characterise the effects of these cigarette substitutes on human host defence mechanisms. Accordingly, the current study compared the effects of the extracts of a basic test EC aerosol, with and without nicotine, with those of cigarette smoke, on several functions of human neutrophils and indices of platelet activation in vitro.
Our findings showed that the ECEs suppressed the immune-protective activities of neutrophils, specifically ROS production and elastase release by activated cells, which was not augmented by inclusion of nicotine. In this context, the neutrophil-suppressive effects of the ECEs appeared to be mediated by toxicants other than nicotine, most likely the tobacco flavourant and/or thermal breakdown products of the propylene glycol and glycerine constituents of the basic EC liquid. This may relate to the contention that nicotine, unlike acrolein and acetaldehyde, appears to be primarily addictive as opposed to being cytotoxic [23].
Not surprisingly, CSE was found to be more suppressive than the test ECEs with respect to effects on neutrophil ROS generation and elastase release. In addition, CSE, but not the test ECEs, attenuated NET formation. With respect to their role in host defence, the primary activity of NETs appears to be immobilisation of microbial pathogens, enabling exposure to bactericidal ROS and proteolytic enzymes in particular, both of which are key anti-infective protective mechanisms of neutrophils. In this context, the apparent sparing of NETosis by the test ECEs indicates that their predominant inhibitory effects on neutrophil reactivity relate to suppression of the generation of ROS and release of elastase. The absence of effects of the two test ECEs on NETosis may result from the failure of these extracts to achieve a level of inhibition of ROS production necessary to attenuate NET formation. Alternatively, albeit unexplored in the current study, the test ECEs may have more subtle qualitative, as opposed to quantitative, effects on NETs, such as promoting alterations in the structure, composition and function of these complex extracellular DNA–histone/protein scaffolds. This issue should be explored in future studies.
Relative to the effects of the test ECEs, the more potent inhibitory actions of CSE on neutrophils are most likely due to the fact cigarette smoke contains substantially more toxicants than ECEs. Nevertheless, despite having less toxicants than CSE, the test ECEs still had significant suppressive effects on the antimicrobial potential of neutrophils, indicating that they are not completely risk free. This contention is supported by the findings of Corriden et al. [24] who found that exposure to ECE resulted in impairment of neutrophil-protective functions such as chemotaxis, ROS production, NETosis and phagocytosis of bacterial bioparticles. From their data, the authors proposed that ECE-exposed neutrophils are less capable of performing their antimicrobial functions [24]. Indeed, in a mouse model of ECE exposure, the treated mice had increased bacterial burdens compared with controls [24]. In contrast to this study, several in vitro and in vivo studies found that exposure of neutrophils to ECE, as well as ECE use, caused activation of the pro-inflammatory activities of neutrophils, e.g. upregulation of matrix metalloproteinase-9 and elastase secretion/activity [25–27]. These disparities may be explained by differences in the methods used between these studies and the current study or by other confounding factors such as differences in the models of exposure used.
With respect to the effects of CSE on human neutrophils, our findings are consistent with the study reported by Bokaba et al. [14], which found that CSC attenuated NET formation and oxygen consumption (as an indirect measure of ROS production) by PMA-activated human neutrophils in vitro. Earlier studies (cited by Matthews et al. [28]) also found that in vitro exposure of neutrophils to cigarette smoke or CSE inhibited PMA-induced ROS generation, although effects on unstimulated cells were contradictory. Another study showed that low levels of CSE (1%) caused a reduction in the ability of neutrophils to mount a respiratory burst against bacterial stimuli [28]. Possible implications of these results and those of the current study are that cigarette smoking may lead to a reduction in the ability of neutrophils to eliminate microbial pathogens.
In the case of platelets, these cells not only play an important role in haemostasis, but also contribute to host antimicrobial defences by facilitating the recruitment of neutrophils to sites of inflammation and infection, promoting both adhesion to vascular endothelium and effector cell functions [29, 30]. In the current study, CSE, but not the test ECEs, was shown to inhibit the expression of the key platelet pro-adhesive, activation marker CD62P on ADP-activated platelets. In a manner similar to the effects of the test ECEs on neutrophils, the inhibitory effects seen with CSE are likely to be related to the higher concentration of toxicants in cigarette smoke. Nevertheless, although the test ECEs were without significant effects on ADP-mediated upregulation of CD62P, we cannot exclude possible modulation of platelet function activated by alternative stimuli, such as collagen, thrombin or thromboxane A2, as well as effects on other types of platelet receptor. With respect to the observed absence of effects of the two test ECEs on NP aggregation, this is most probably underpinned by the failure of these extracts to inhibit expression of CD62P. However, the lack of significant effects of CSE on NP aggregation is more difficult to explain given the potent inhibitory effects of this extract on upregulation of ADP-mediated expression of the adhesion molecule. This may relate to a requirement for increased concentrations of CSE and/or extended incubation times to detect effects on NP interactions.
Limitations of our study include the in vitro design; the use of blood neutrophils as opposed to cells derived from body fluids such as sputum; the short exposure times of neutrophils and platelets to the ECEs in particular; and the fact that our test ECEs may not be entirely representative of the various EC commercial brands that contain different constituents at varying concentrations.
In conclusion, our results show that the EC aerosol extracts tested in the current study adversely affect cells of the human innate immune system, albeit to a lesser extent than those of CSE. Nevertheless, if operative in the airways, these observed inhibitory effects on activation of neutrophil antimicrobial mechanisms may predispose EC users for development of bacterial infections of the airways. In this context, the association of cigarette smoking with increased susceptibility for development of respiratory infections is well established [31]. However, to the best of our knowledge, a similar, compelling link with EC usage and increased risk of infection in humans has not yet been recognised. Given the implications for public health, more intensive studies, particularly in the clinical setting, are clearly necessary to fully elucidate the anti-infective toxicities associated with the use of these products.
Footnotes
Provenance: Submitted article, peer reviewed.
Conflict of interest: None declared.
- Received September 27, 2022.
- Accepted March 17, 2023.
- Copyright ©The authors 2023
This version is distributed under the terms of the Creative Commons Attribution Non-Commercial Licence 4.0. For commercial reproduction rights and permissions contact permissions{at}ersnet.org
References










