



Lack of evidence for the involvement of Merkel cell polyomavirus in pulmonary Langerhans cell histiocytosis
- Fanélie Jouenne1,2,
- Jérôme Le Goff1,3,
- Emmanuelle Bugnet4,
- Maud Salmona1,3,
- Véronique Meignin5,
- Gwenaël Lorillon4,
- Aurélie Sadoux2,
- Janine Cherot3,
- Céleste Lebbé1,6,
- Samia Mourah1,2 and
- Abdellatif Tazi1,4⇑
- 1Université de Paris, INSERM U976, Paris, France
- 2Pharmacogenomics Dept, Hôpital Saint-Louis, Assistance Publique-Hôpitaux de Paris, Paris, France
- 3Infectious Agents Department, Virology Unit, Hôpital Saint-Louis, Assistance Publique-Hôpitaux de Paris, Paris, France
- 4National Reference Centre for Histiocytoses, Pulmonology Dept, Hôpital Saint-Louis, Assistance Publique-Hôpitaux de Paris, Paris, France
- 5Pathology Dept, Hôpital Saint-Louis, Assistance Publique-Hôpitaux de Paris, INSERM, UMR_S1165, Paris, France
- 6Dermatology Dept, Hôpital Saint-Louis, Assistance Publique-Hôpitaux de Paris, Paris, France
- Abdellatif Tazi, Service de Pneumologie, Hôpital Saint-Louis, 1 Avenue Claude Vellefaux, 75475, Paris cedex 10, France. E-mail: abdellatif.tazi{at}aphp.fr
Abstract
Compared to control lung tissues from smokers, MCPyV DNA is rarely detected in PLCH lesions and is not associated with alterations of the MAPK pathway. A viral trigger in PLCH pathogenesis remains elusive. https://bit.ly/2xKmkIo
To the Editor:
Langerhans cell histiocytosis (LCH) is currently considered a rare neoplastic disease with an inflammatory component, driven by genomic alterations in the mitogen-activating protein kinase (MAPK) pathway, including BRAFV600E, MAP2K1 mutations and BRAF deletions [1, 2]. Lung involvement is frequently observed in adult LCH patients, almost exclusively in young smokers of both sexes [3]. The aetiology of LCH, including pulmonary LCH (PLCH), remains unknown. A viral triggering factor of LCH has long been suspected, but the results of studies are inconclusive [4, 5].
Recently, Murakami et al. [6] reported the detection of increased numbers of Merkel cell polyomavirus (MCPyV) DNA sequences in nonpulmonary LCH tissues compared to control tissues from Japanese patients and suggested an association of MCPyV with the presence of the BRAFV600E mutation in LCH lesions [7]. MCPyV is composed of the regulatory proteins large T (LT) antigen (Ag) and small T (ST)-Ag, and structural proteins VP1 and VP2 [8]. MCPyV is known to be involved in the rare aggressive human skin cancer Merkel cell carcinoma (MCC), 80% of MCC tumours being positive for the MCPyV genome [8].
Our purpose was to investigate the potential involvement of MCPyV in PLCH. We used highly sensitive real-time PCR and immunohistochemistry (IHC) to detect the presence of MCPyV DNA sequences and LT-Ag in PLCH biopsies compared to that in control lung tissues. We also searched for an association between the presence of MCPyV DNA and MAPK mutation status of PLCH lesions.
Nine PLCH patients (eight with single-system disease; five females; mean age 47 years, range 30–61 years; all smokers) and 19 controls (14 females; mean age 65 years, range 28–86 years; nine smokers and 10 nonsmokers) were included. Grossly normal lung tissue was obtained at the time of thoracic surgery for localised lung disease (table 1). The study was performed in accordance with the Helsinki Declaration and was approved by the INSERM Institutional Review Board and Ethics Committee in Paris, France (IRB number 13-130). All patients gave informed consent for the use of their information for research.
Main clinical characteristics of the nine patients with pulmonary Langerhans cell histiocytosis (PLCH) and the 19 lung control tissue patients as well as detection of Merkel cell polyoma virus by real-time PCR
Formalin-fixed paraffin-embedded (FFPE) PLCH biopsies were immunostained with an anti-CD1a antibody (clone 010; Dako, Waldbronn, Germany) to identify lesional areas that were macrodissected for molecular biology analysis. FFPE control lung tissues were optically normal or displayed mild changes associated with smoking. We also performed IHC staining for the detection of MCPyV in PLCH and control lung lesions, using an antibody directed against MCPyV LT-Ag (clone CM2B4; Santa Cruz Biotechnology, Dallas, TX, USA). Immunostaining was performed with a BenchMark Ultra automated immunostainer (Roche-Ventana, Basel, Switzerland) according to the manufacturer's instructions.
DNA was extracted as previously described [9]. MCPyV DNA was detected by real-time PCR assays targeting genes coding for VP1, ST and LT using specific primers and probes [8]. Real-time DNA amplification was performed on an ABI PRISM 7500 SDS thermocycler (Thermo Fisher Scientific, Waltham, MA, USA). The results are reported as a qualitative score as described in table 1. DNA from skin biopsies of three MCC lesions was used as a positive control. Fisher's exact test was performed to compare categorical variables between groups of patients, with p-values <0.05 denoting statistical significance. To estimate the viral load of the MCPyV genome in lung biopsy samples, we used standard curves established for each target (VP1, ST and LT) with a plasmid including the full genome of MCPyV (RepMCV-R17a; Addgene, Watertown, MA, USA), using real-time PCR [10].
A custom-designed next-generation sequencing panel of 74 genes, including genes involved in the MAPK pathway, was used for genotyping PLCH tissue biopsies [2]. The exon 12 BRAF deletion was assessed using a custom pyrosequencing assay on a PyroMark Q48 instrument (Qiagen, Hilden, Germany) [2].
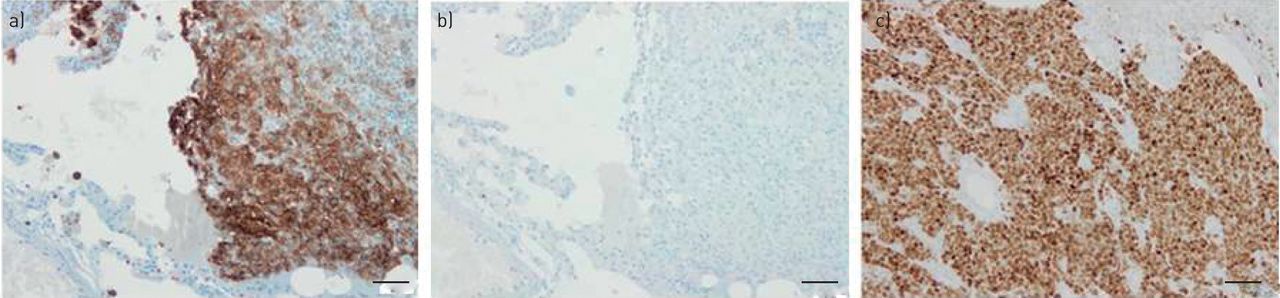
Using real-time PCR, the three skin MCC biopsies were positive for MCPyV genes (table 1). VP1, ST and LT genes were found positive in, respectively, 10, 10 and eight lung specimens tested (table 1). Three samples (one PLCH and two nonsmoker control lung tissues) were positive for all targets, whereas 10 specimens (six PLCH) were negative for all targets. Four lung tissues (one PLCH) were positive for two targets and the 11 remaining samples (one PLCH) expressed only one MCPyV DNA target. Taken together, three (33%) out of nine PLCH lesions, eight (89%) out of nine smoker control tissues and seven (70%) out of 10 nonsmoker control tissues were positive for at least one MCPyV DNA target. Compared to positive skin MCC biopsies, IHC staining for MCPyV LT-Ag was consistently negative in the seven available PLCH lesions as well as smoker or nonsmoker controls tissues (figure 1).

{kind=link}
Immunohistochemistry. a) CD1a immunostaining of a lung biopsy from a patient with pulmonary Langerhans cell histiocytosis showing accumulation of large numbers of positive cells. b) Merkel cell polyomavirus large T antigen immunostaining on a serial section of the same lung biopsy showing that no positive cells were identified. c) Merkel cell polyomavirus large T antigen antibody immunostaining of a skin biopsy from a patient with cutaneous Merkel cell carcinoma demonstrating intensely positive tumour cells. Original magnification ×200. Scale bars=50 μm.
The proportion of MCPyV DNA detected in PLCH biopsies was significantly lower than that detected in control tissues from both smokers and nonsmokers (p=0.035, Fisher's exact test comparing PLCH and control specimens). Median viral load ratios of the three genes VP1, ST and LT were not different comparing PLCH, smoker and nonsmoker lung tissues (Kruskal–Wallis, p=0.747).
An alteration of the MAPK pathway was identified in six (67%) out of nine PLCH lesions, including four BRAFN486_P490del deletions and two MAP2K1 mutations (R108W and G128D). Only one of these lesions harboured MCPyV DNA.
Our results are inconsistent with the previous study from the only group reporting on the presence of MCPyV DNA sequences in LCH lesions. The authors reported that 12 (92%) out of 13 nonpulmonary LCH lesions were positive for MCPyV DNA [6], whereas in our study, three (33%) out of nine of PLCH lesions contained DNA for at least one MCPyV target. Furthermore, based on the same MCPyV DNA detection technique and using the same criteria as Murakami et al. [6], i.e. the presence of the LT target, only one PLCH lesion was positive in our study.
A main difference between PLCH and other clinical forms of LCH is that the former is strongly associated with tobacco smoking [3]. Strikingly, we found a lower rate of detection of MCPyV DNA in PLCH lesions compared to that in smoking control lung tissues. Murakami et al. [6] found similar numbers of single-system and multisystem nonpulmonary LCH lesions harbouring MCPyV DNA. Thus, the fact that our series comprised almost exclusively single-system PLCH probably does not explain the discrepancy between the two studies. Furthermore, we could not confirm that the detection of MCPyV DNA was linked to the MAPK status of LCH lesions.
A possible explanation of these discordant results could be linked to ethnicity difference. Indeed, MCPyV epidemiology varies according to the ethnicity and geographic location of the subjects studied [11]. Consistently, discordant results for pathogen detection have been previously reported between Japanese and Caucasian patients in different diseases [12, 13].
The presence of MCPyV DNA in tested control lung tissues, regardless of the smoking status, is in accordance with what has been previously reported, although at a lower proportion (10–50%), considering either LT only [14], or VP1 and LT targets [15, 16]. The fact that we also tested for the ST target probably explains the higher rate of positive control lung tissue samples in our series.
In summary, our results do not support the involvement of MCPyV in the pathogenesis of PLCH. Additional studies that include patients of different ethnicities, geographical areas and LCH subtypes are needed to further clarify the potential involvement of MCPyV in LCH.
Acknowledgements
The authors thank S. Mercier-Delarue for performing plasmid experiments (Infectious Agents Dept, Virology Unit, Hôpital Saint-Louis, Assistance Publique-Hôpitaux de Paris, Paris, France) and M. Mao (Pulmonology Dept, Hôpital Saint-Louis, Assistance Publique-Hôpitaux de Paris, Paris, France) for her technical assistance.
Footnotes
Conflict of interest: F. Jouenne has nothing to disclose.
Conflict of interest: J. Le Goff has nothing to disclose.
Conflict of interest: E. Bugnet has nothing to disclose.
Conflict of interest: M. Salmona has nothing to disclose.
Conflict of interest: V. Meignin has nothing to disclose.
Conflict of interest: G. Lorillon reports support for travel and accommodation from Vitalaire and Chiesi outside the submitted work.
Conflict of interest: A. Sadoux has nothing to disclose.
Conflict of interest: J. Cherot has nothing to disclose.
Conflict of interest: C. Lebbé reports honoraria from, and consultancy, and serving on a speakers bureau and an advisory board for Amgen; grants and honoraria from, and consultancy, and serving on a speakers bureau and an advisory board for BMS; grants and honoraria from, and consultancy, and service in an advisory role and on an advisory board for, and travel and accomodation for meetings from MSD; grants and honoraria, and consultancy, and service in a speakers bureau, in an advisory role and on an advisory board for Roche and Novartis; consultancy for and honoraria from Pierre Fabre; consultancy for Sanofi and Merck Serono; and honoraria from Pfizer and Incyte, all outside the submitted work.
Conflict of interest: S. Mourah reports consulting for Novartis and Roche outside the submitted work.
Conflict of interest: A. Tazi reports personal fees from Bristol-Myers Squibb for speaking at a conference, and travel and accommodation support from Boehringer Ingelheim, Teva, Vitalaire and AstraZeneca, all outside the submitted work.
- Received September 4, 2019.
- Accepted March 24, 2020.
- Copyright ©ERS 2020
This article is open access and distributed under the terms of the Creative Commons Attribution Non-Commercial Licence 4.0.