Article Text

Abstract
Background: Primary ciliary dyskinesia (PCD) is a severe inherited disorder characterised by chronic respiratory disease, male infertility, and, in ∼50% of affected individuals, a left-right asymmetry defect called situs inversus. PCD is caused by defects in substructures of the ciliary and flagellar axoneme, most commonly loss of the outer dynein arms. Although PCD is believed to involve mutations in many genes, only three have been identified.
Methods: To facilitate discovery of new PCD genes, we have used database searching and analysis to systematically identify the human homologues of proteins associated with the Chlamydomonas reinhardtii outer dynein arm, the best characterised outer arm of any species.
Results: We find that 12 out of 14 known Chlamydomonas outer arm subunits have one or more likely orthologues in humans. The results predict a total of 24 human genes likely to encode outer dynein arm subunits and associated proteins possibly necessary for outer arm assembly, plus 12 additional closely related human genes likely to encode inner dynein arm subunits.
Conclusion: These genes, which have been located on the human chromosomes for easy comparison with known or suspected PCD loci, are excellent candidates for screening for disease-causing mutations in PCD patients with outer and/or inner dynein arm defects.
- AK, adenylate kinase
- DC, docking complex
- HC, heavy chains, IC, intermediate chains, LC, light chains
- PCD, primary ciliary dyskinesia
- Chlamydomonas
- cilia
- dynein
- flagella
- immotile cilia syndrome
- Kartagener’s syndrome
- primary ciliary dyskinesia
Statistics from Altmetric.com
- AK, adenylate kinase
- DC, docking complex
- HC, heavy chains, IC, intermediate chains, LC, light chains
- PCD, primary ciliary dyskinesia
- Chlamydomonas
- cilia
- dynein
- flagella
- immotile cilia syndrome
- Kartagener’s syndrome
- primary ciliary dyskinesia
Primary ciliary dyskinesia (PCD; MIM 242650), also known as immotile cilia syndrome, is the name given to a group of genetic disorders of motile cilia that includes Kartagener syndrome (MIM 244400).1 The disease, which affects about one in 20 000 humans,2 usually involves a defect in a substructure of the “9+2” axonemes of the cilia and flagella (fig 1): patients have been reported with loss of the outer dynein arms, the inner dynein arms, the outer arms and the inner arms, the radial spokes, and the central pair of microtubules. As a result, ciliary and flagellar motility is abnormal, leading to a similar clinical profile that includes chronic or recurrent sinopulmonary infections, infertility in males, and a reversal in the placement of internal organs in about half of affected individuals. PCD is genetically heterogeneous and is likely caused by defects in many different genes; however, to date only three PCD genes have been identified.3–5
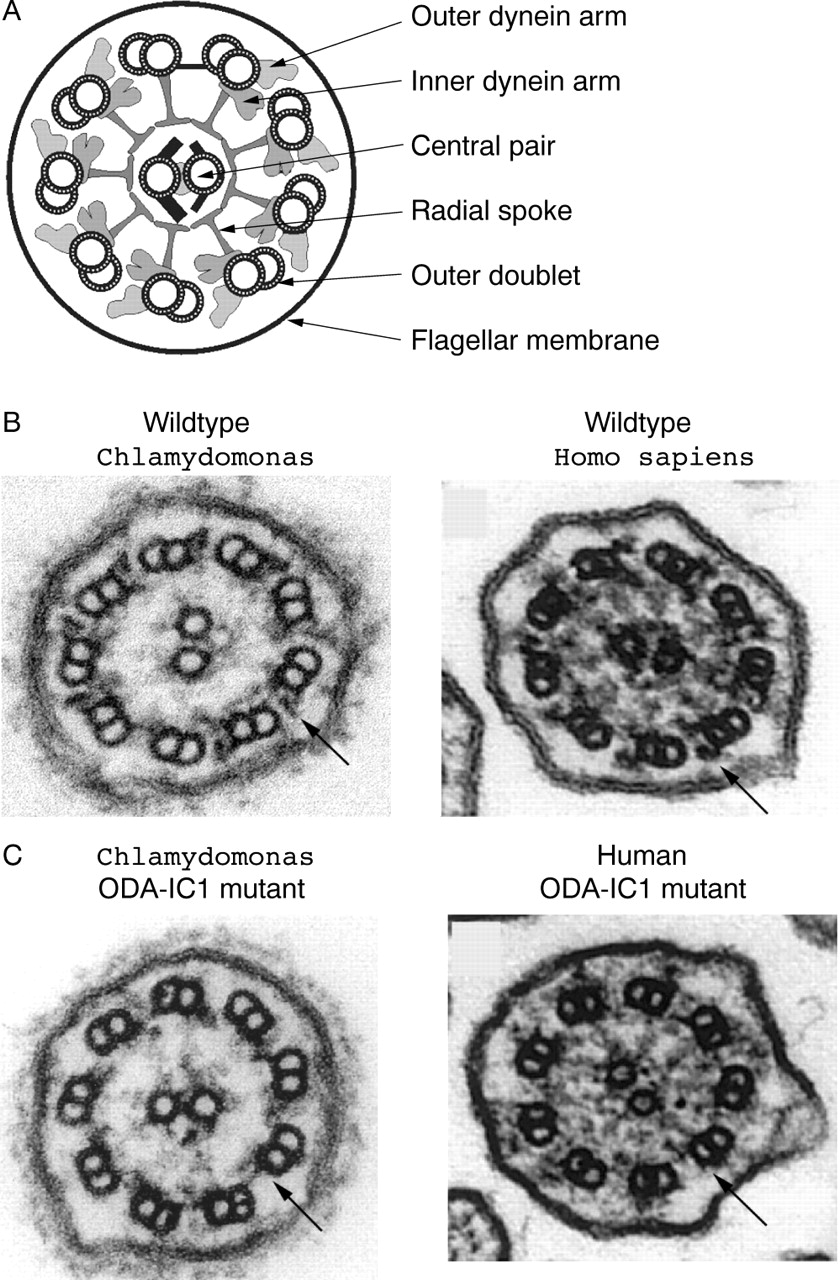
The “9+2” axoneme is structurally similar in Chlamydomonas and humans. (A) Schematic cross section of a 9+2 cilium showing the major axonemal substructures. (B, C) Cross sections of wildtype (B) and mutant (C) cilia from Chlamydomonas and humans. An outer dynein arm in each wildtype cilium is marked with an arrow. In the mutants of both species, a defect in the gene encoding the IC1 subunit of the outer arm dynein results in loss of the outer arms (arrows). The Chlamydomonas images are from Wilkerson et al,24 and the human images are from Pennarun et al3 and are used with permission.
The discovery of additional genes involved in PCD would be greatly facilitated by more complete information on the human genes that encode axonemal structures. Proteomic analysis of isolated ciliary axonemes from primary cultures of human bronchial epithelial cells has resulted in a list of over 200 potential axonemal proteins,6 but identification of the defective gene in a patient with PCD would be much simplified by knowledge of the subset of genes necessary for assembly of the specific axonemal substructure that is defective in the patient. Currently, data on the proteins that make up each of the substructures of the axoneme are most complete for the unicellular green alga Chlamydomonas reinhardtii, which has been used extensively to study the assembly, composition, and mechanism of action of cilia and flagella. The Chlamydomonas cell has two flagella extending from its anterior end that it uses for swimming. Structurally, these flagella are virtually indistinguishable from human airway cilia (fig 1), and biochemical and proteomic studies indicate that most of the axonemal proteins have been highly conserved throughout evolution,7 so that their human orthologues can be readily identified. Moreover, mutations in Chlamydomonas are known that cause loss of each of the major axonemal substructures, thus mimicking nearly all of the ultrastructural defects seen in PCD. As a result, Chlamydomonas is an excellent model system for PCD, and information on the genes necessary for assembly of an axonemal structure in Chlamydomonas should be directly transferable to humans by identification of the human orthologues of these genes. Indeed, such information provided the basis for discovery, using a candidate gene approach, of the first two PCD genes.3,4
The most frequent cause of PCD is loss of the outer dynein arms,8–10 and in this study we use known Chlamydomonas genes involved in assembly of the outer dynein arms to identify their likely human counterparts. Outer dynein arms are tightly bound to one outer doublet microtubule and use the energy of ATP hydrolysis to make transient interactions with an adjacent doublet microtubule to produce force for ciliary beating. The Chlamydomonas outer dynein arm is a 1.2 MDa complex composed of three heavy chains (HC), two intermediate chains (IC), and at least nine light chains (LC) (see fig 3). The HCs are the sites of the ATP hydrolysis required for ciliary motility. The ICs bind to the HCs and appear to be involved in regulation of HC activity and attachment of the dynein to the microtubule. The nine LCs are a diverse group of proteins including a Ca2+ binding protein, thioredoxin-like molecules, and an extremely highly conserved peptide that is also found in several other motor and non-motor complexes. The outer arm is bound to the doublet microtubule by a distinct structure termed the docking complex (DC), which is essential for outer arm assembly and contains two coiled-coil proteins and an EF-hand protein similar to calmodulin. An additional, less well characterised complex that includes a protein called ODA5 and an associated adenylate kinase (AK) also is essential for outer arm assembly. All of these subunits have been cloned and sequenced (table 1). Importantly, mutational analysis indicates that defects in most of the above proteins, even including the LCs, can cause loss of the outer dynein arms in Chlamydomonas (table 1). Therefore, the human genes encoding orthologues of all of these proteins are candidates for causing PCD in those human patients with outer dynein arm defects. In addition to identifying these potential orthologues, we also place the outer dynein arm loci on the human genetic map for easy comparison with known or suspected PCD loci.
Known Chlamydomonas outer dynein arm subunits and associated proteins
The nomenclature of dynein genes is complicated and convoluted with multiple names given to the same sequence and the same names given to different sequences. This has produced a situation where it is difficult for even an expert in the field to follow the literature. To overcome this problem, we have used the official names given to the genes by the HUGO Gene Nomenclature Committee (http://www.gene.ucl.ac.uk/nomenclature/) and provided RefSeq numbers for the proteins (in the text) and corresponding mRNAs (in the tables) whenever possible. We have also extensively documented GenBank accession numbers (acc. no.) and NCBI GeneID tags throughout the document. Since even this may still leave uncertainty, we have also included the predicted peptide sequence encoded by each of the loci in the supplemental material (available at http://www.jmedgenet.com/supplemental).
METHODS
Bioinformatics
BLAST searches were performed using the NCBI BLAST server (http://www.ncbi.nlm.nih.gov/BLAST/). Multiple sequences were aligned with ClustalW11 and phylogenetic trees were drawn with NJPlot.12 Human genome positions were identified using the NCBI human genome BLAST server (http://www.ncbi.nlm.nih.gov/genome/seq/HsBlast.html) and the Golden Path web server at the University of California at Santa Cruz (http://genome.ucsc.edu).
RESULTS
HC genes
Dynein HCs are large ∼450 kDa proteins that convert the energy of ATP to force. These proteins have an N-terminal tail connected to a C-terminal globular head. The N-terminal tail binds ICs and most LCs and is thought to be involved in binding the dynein motor to microtubules or cargo in an ATP insensitive manner. The globular head is made up of six AAA domains organised in a hexameric ring with a short extension protruding from the ring that is postulated to bind microtubules in an ATP sensitive manner.13 The Chlamydomonas outer dynein arm is composed of three HCs termed DHCα (Q39610), DHCβ (Q39565), and DHCγ (Q39575). Mutations in the ODA4 and ODA2 genes, which encode DHCβ and DHCγ, respectively, block assembly of the entire outer arm complex and cause an approximately two thirds reduction in swimming speed.14–17 Cells carrying a mutation in ODA11, which encodes DHCα, assemble an outer dynein arm that lacks DHCα and the DHCα associated LC5. These cells swim only slightly slower than normal.18 The number of HCs in flagellar outer arm dyneins varies with species. Like Chlamydomonas, Tetrahymena has three distinct HCs, while sea urchins have two HCs.19 The organisation of human outer arm dynein has not been examined, but in two other vertebrates, trout and pigs, the outer arm appears to be composed of two HCs.20,21
The human dynein HC genes were identified by BLAST searching the non-redundant database with protein and nucleotide sequence from the region surrounding the first nucleotide binding P-loop of dynein. This region has been cloned from many species and is adequate to identify different dynein isotypes. Searches were initiated using Chlamydomonas ODA-DHCβ and repeated using sequences identified by this search until no new sequences were identified. Sixty three supposedly human dynein HC sequences were found. This list is highly redundant, as multiple laboratories have identified many of the dynein HC sequences independently.
Each of the 63 sequences was used to search the human genomic sequence using the NCBI human genome BLAST server. All the sequences except four (Z83804, Z83799, U61737, and Z83803) could be located in the human genomic sequence. These four are also missing from the Celera public human genome sequence, suggesting that they may not be of human origin. Supporting this idea, Z83804 and Z83799 are similar to mouse sequences, U61737 is similar to a rat sequence, and Z83803 is similar to a Drosophila hydei sequence.
The remaining 59 sequences aligned to 15 distinct genomic loci (tables 2–4). No additional dynein HC-like sequences were found in the genome. Some of the dynein cDNA sequences showed small polymorphisms with the genomic sequence. These probably reflect variation within the human population, or errors generated by PCR amplification of the cDNA when the HCs were originally cloned.
Human outer dynein arm heavy chain loci
Sequences corresponding to the highly conserved dynein HC domain GPAGTGKT…FITMNP encoded by each of the human genomic loci and by each of the known Chlamydomonas dynein HC genes were aligned by ClustalW. If the published peptides did not span the sequence, the sequence was extended using genomic sequence from the region to predict the missing amino acids. The alignment was used to draw a phylogenetic tree (fig 2). There are five human genes that encode proteins similar to Chlamydomonas DHCβ and DHCγ (fig 2, table 2). Three of these proteins, DNAH11 (NP_003768), DNAH17 (NP_003718), and DNAH9 (NP_001363), encoded at 7p21, 17q25, and 17p12, are most similar to Chlamydomonas DHCβ.17 DNAH5 (NP_001360) and DNAH8 (NP_001362), from 5p15 and 6p21, are most similar to Chlamydomonas DHCγ.16 Humans do not appear to have an orthologue of Chlamydomonas DHCα.

Phylogenetic tree showing the relationship between the dynein HCs of Chlamydomonas and humans. An ∼100 amino acid peptide spanning the region from GPAG to FITMNP near the first P-loop of each of the dyneins was aligned by ClustalW and developed into a phylogenetic tree. The numbers show the bootstrap values from 1000 iterations. The outer dynein arm γ and β branches of the tree are marked near the bottom of the tree.
Eight human genes encode inner arm dynein HCs (table 3) and are tabulated here because they are candidates for causing PCD in patients with defects in the inner dynein arm. Among the inner arm dynein HCs, DNAH10 and DNAH2 correspond to DHC1α and DHC1β of the Chlamydomonas inner arm dynein 1. This inner arm also contains the ICs IC140 and IC138 (see below) as well as at least two LCs.
Human inner dynein arm heavy chain loci
A single human gene encodes each of the cytoplasmic dynein HCs DHC1 and DHC2 (table 4). DHC1 is the conventional cytoplasmic dynein HC isoform and corresponds to DHC1a in Chlamydomonas. DHC2, the orthologue of Chlamydomonas DHC1b, is involved in the process of intraflagellar transport, which is necessary for assembly of cilia and flagella.
Human cytoplasmic dynein heavy chain loci
IC genes
The outer dynein arm ICs are WD-repeat proteins that are part of a family of proteins that includes cytoplasmic and inner dynein arm ICs (table 5). Chlamydomonas IC1 (formerly IC78 or IC80) (Q39578) binds to α-tubulin and is thought to be involved in binding the base of the outer dynein arm to the flagellar microtubule.22Chlamydomonas IC2 (formerly IC69 or IC70) (P27766) is thought to be involved in regulation of the motor activity of the outer dynein arm.23 Defects in the ODA9 and ODA6 genes, which encode Chlamydomonas IC1 and IC2, respectively, block assembly of the outer dynein arm; as a result, the cells swim at about one third normal speed.15,23,24
Human dynein intermediate chain genes
Pennarun et al3 identified DNAI1 (NP_036276) as the human orthologue of Chlamydomonas IC1 (Q39578). The Chlamydomonas and human proteins are 43% identical and have a BLAST E value of 1e-150. Pennarun et al25 and Bartoloni et al26 identified DNAI2 (NP_075462) as the human orthologue of Chlamydomonas IC2 (P27766). The Chlamydomonas and human proteins are 48% identical and the comparison has a BLAST E value of 1e-154.
Searching the human genomic sequence with Chlamydomonas IC1 and IC2, or human DNAI1 and DNAI2 identified four regions of significant homology on chromosomes 1, 9, 10, and 17. Reiteratively searching identified additional loci on chromosomes 1, 2, and 7. The locus at chromosome 9p21-p13 encodes outer arm IC DNAI1, while the locus at 17q25 encodes outer arm IC DNAI2. The loci at 7q21 and 2q31 encode cytoplasmic dynein ICs DNCI1 and DNCI2, respectively. The locus at chromosome 10q11 encodes a relatively uncharacterised region of sequence (XM_208483) that at the peptide level is almost identical to cytoplasmic dynein intermediate chain DNCI2. This locus is likely a pseudogene derived from integration of a DNCI2 cDNA as it is almost continuous with the DNCI2 cDNA sequence and has a stop codon within the predicted coding region. The locus at chromosome 1p31 encodes a novel protein that is similar to dynein ICs. There are two cDNA clones from this region: AK026782 appears to be full length while AL133617 is from the 3′ end of the gene. The peptide encoded by AL133617 should be included within the protein encoded by the longer AK026782 cDNA but is not due to a frame shift in AK026782. This protein has now been curated as WDR78. The locus at 1p22 encodes a testis expressed protein called WDR63.
Phylogenetic analysis (not shown) supports the published assignment of DNAI1 and DNAI2 as outer arm dynein ICs and DNCI1 and DNCI2 as cytoplasmic dynein ICs. The presumed pseudogene XM_208483 also groups with the cytoplasmic dynein ICs. The remaining two human sequences, WDR63 and WDR78, group with Chlamydomonas inner dynein arm IC140 (AAD45352) and IC138 (AAU93505), suggesting that they are subunits of the inner dynein arm.
LC genes
LC1
Chlamydomonas outer dynein arm LC1 (AAD41040) is a leucine rich repeat protein. No mutations in the gene encoding LC1 are known to exist in Chlamydomonas, so the importance of this LC to arm structure is unknown.
Benashski and colleagues27 identified EST AA923426 as a potential human orthologue. This EST has now been curated as a hypothetical protein C14orf168 (table 6). Searching the human genomic sequence with Chlamydomonas LC1 or human C14orf168 identified three regions of homology on chromosomes 14, 2, and 12. The locus at 14q24 encodes C14orf168. The locus on chromosome 2 (Entrez Gene reports the locus at 15q26) encodes a small nuclear ribonucleoprotein polypeptide A′ (NP_003081), whereas the locus at 12q21 encodes a glycoprotein called lumican (NP_002336). Chlamydomonas LC1 and human C14orf168 are 55% identical and have BLAST E values of 3e-38, supporting the idea that they are orthologues. The BLAST E values for comparisons between Chlamydomonas LC1 and the ribonucleoprotein or lumican are relatively poor (3e-06 and 7e-05), indicating that they are not likely to be orthologues.
Human dynein light chain 1 loci
LC2
Chlamydomonas LC2 (AAB58383) is homologous to mouse TCTEX2 and is very distantly related to TCTEX1.28 Murine TCTEX1 and TCTEX2 are encoded within the t complex. This region of chromosome 17 is involved in an extreme form of meiotic drive whereby in +/t males the t version of chromosome 17 is transmitted to >90% of the offspring. The meiotic drive is thought to be the result of differential phosphorylation of flagellar dynein affecting sperm motility.28 Mutations in oda12, which encodes LC2 in Chlamydomonas, block outer dynein arm assembly.29 A second Chlamydomonas LC2-like protein called Tctex2b (DAA05278) was recently found associated with the inner dynein arm. Mutant analysis indicated that the protein is not required for inner arm assembly but is required for arm function as cells lacking this protein swam more slowly than normal.30
Searching the human genome database with Chlamydomonas LC2 identified two regions of homology on chromosome 1 and additional regions of homology on chromosomes 3 and 6 (table 7). The loci at 1p31 and 3q29 encode poorly characterised proteins called FLJ40873 (NP_689878) and MGC33212 (NP_689986), while the locus at 6q27 encodes a protein called TCTE3 (NP_777570). The locus at 1p34.1 has been predicted to encode an uncharacterised protein called LOC343521 (NP_001013654). This locus and the other three appear to be expressed, as there are EST sequences corresponding to all four loci in the databank. These four proteins group phylogenetically with Chlamydomonas LC2 and Tctex2b, as well as with a sea urchin homologue of LC2 (BAA24185) (data not shown). TCTE3 groups closely with Chlamydomonas LC2 and thus is assigned to the outer dynein arm; MGC33212 groups closely with the Chlamydomonas Tctex2b and thus is assigned to the inner dynein arm. The trees do not definitively group FLJ40873 and LOC343521 with either the outer or the inner dynein arm LC2s; nevertheless, they should be considered candidates for PCD genes that map to chromosome 1p31-34.
Human dynein light chain 2 loci
Tctex1 is not found in the Chlamydomonas flagellar outer dynein arm but is found in the inner dynein arm of the Chlamydomonas flagellum31 and the outer dynein arm of the sea urchin flagellum.32 It is not known if TCTEX1 is a subunit of human outer arm dynein, but since it is found in the sea urchin outer dynein arm, database searches were carried out to identify the human homologues. Six Tctex1-like sequences are encoded by the human genome on chromosomes X, 6, 17, 4, 20, and 2 (table 7). The loci at 6q25 and Xp21 encode proteins TCTEL1 (NP_006510) and TCTE1L (NP_006511), respectively. The gene encoding the latter protein was considered a candidate for the retinitis pigmentosa 3 gene and is sometimes referred to as RP3. However, it is now known that the real RP3 gene encodes the retinitis pigmentosa GTPase regulator.33 The other four genomic loci do not appear to be expressed at a significant level, as these sequences could not be found in the protein or EST databanks. The locus on chromosome 17 is likely to be a pseudogene as there are stop codons within the predicted coding region.
LC3 and LC5
Chlamydomonas LC3 (Q39592) and LC5 (AAB03681) are members of the thioredoxin family.34 Searching the databanks with these proteins identified the sea urchin outer dynein arm IC1 protein (BAA09934), Ciona dynein intermediate chain 3 (BAB68388), human TXNDC3 (also known as NM23-H8) (NP_057700), human TXNDC6 (also known as thioredoxin-like 2 TXL-2) (NP_835231), and a large number of thioredoxin-like proteins. The sea urchin and Ciona dynein ICs and the two human proteins are composed of both thioredoxin-like domains and a nucleoside diphosphate kinase-like domain.35,36 No mutations in the Chlamydomonas genes encoding LC3 or LC5 have been found so it not known how mutations in these genes will affect assembly of the outer dynein arm.
Searching the human genomic sequence with Chlamydomonas LC3 and LC5 and the LC3/5-like domain of TXNDC3 identified four regions with homology on chromosomes 3, 4, 7, and 18. These four regions were each used to search the protein database to determine which thioredoxin-like protein is encoded at that region. The locus on chromosome 4 encodes a widely expressed isoform of thioredoxin termed TXN (NP_003320), while the locus on chromosome 18 encodes a sperm specific isoform of thioredoxin termed TXNDC2, also known as Sptrx-237 (NP_115619), that has been localised to the fibrous sheath of the sperm flagellum.38 The locus on chromosome 7 encodes TXNDC3 whereas the locus on chromosome 3 encodes TXNDC6, which is expressed in ciliated cells and binds microtubules39 (table 8). Phylogenetic analysis (using the thioredoxin domains) suggests that human TXNDC3 and TXNDC6 are outer dynein arm subunits as only these proteins group with the known Chlamydomonas and sea urchin outer arm polypeptides in a tree containing thioredoxin-like molecules (data not shown).
Dynein light chain 3 and 5 loci
LC4
Chlamydomonas LC4 (Q39584) is a calcium binding EF-hand protein.40 No mutations in the gene encoding LC4 are known in Chlamydomonas.
Searching the human genomic sequence with Chlamydomonas LC4 identified 14 regions of homology (E<0.001). The top two matches are located on chromosomes 18 and 10. The locus on chromosome 18 encodes centrin-1 (NP_004057), while the locus on chromosome 10 encodes calmodulin-3 (NP_005176). It is possible that the human orthologue of LC4 has diverged to the point that it is not readily recognisable above a background of small EF-hand proteins, that the human outer arm lacks an LC4 orthologue, or that centrin-1 or calmodulin-3 perform the functional role of LC4 in the human outer arm.
LC6 and LC8
Chlamydomonas LC6 (Q39579) and LC8 (Q39580) are members of a highly conserved class of dynein LCs. LC8 is one of the most highly conserved proteins known. Chlamydomonas LC8 is 92% identical to human LC8.41 In addition to being a component of the outer dynein arm, LC8 has been found associated with inner arm dynein, myosin V, and cytoplasmic dynein, along with numerous other non-motor proteins including radial spokes, nitric oxide synthetase, and BIM death activator proteins.42 Mutations in the Chlamydomonas FLA14 gene, which encodes LC8, result in multiple flagellar defects, including lack of outer and inner arm dyneins and radial spokes, and loss of retrograde intraflagellar transport.43 Deletion of the gene encoding Chlamydomonas LC6 (ODA13) does not prevent assembly of the outer dynein arm and has very little or no effect on swimming (Pazour and Witman, unpublished results).
Searching the human genomic databank with Chlamydomonas LC8 and LC6 identified two regions of similarity on chromosome 14 and single regions on chromosomes 1, 3, 4, 7, 12, 17, and 22 (table 9). Phylogenetic analysis indicates that DNAL4 (NP_005731) groups with LC6, while all the others group with LC8 (data not shown). Within the LC8 group, DLC2 (NP_542408) is most closely related to Chlamydomonas LC8.
Dynein light chain 6 and 8 loci
Each of the chromosomal regions was searched against the EST databank to determine if the genes were expressed. Three of these loci, 14q31, 4q23, and 3q25, have stop codons within the predicted open reading frames, suggesting that they are pseudogenes. Consistent with this, they are not found in the EST database. Surprisingly, even though the other six sequences are highly conserved, only three of them are represented in the EST databank. These encode DNCL1 (NP_003737) on chromosome 12, DLC2 on chromosome 17, and DNAL4 on chromosome 22. It will be of interest to determine if DLC2 and DNCL1 have similar roles in distinct protein complexes.
LC7
Chlamydomonas LC7a (AAD45881) is a member of the LC7/Roadblock family of proteins.44 Mutations in the Chlamydomonas LC7a gene (ODA15) partially block assembly of the outer dynein arm and cause the cells to swim at about one third normal speed45 (Pazour and Witman, unpublished results). A second LC7/Roadblock family member, named LC7b, was recently found. It appears that both LC7a and LC7b can associate with the outer dynein arm and also with the inner dynein arm I1.46
Two groups of human ESTs (HUM424e02b and AA446298) were previously identified that encoded homologues of LC7/Roadblock; these proteins were named ROBL1 and ROBL2.44 These proteins show differential expression with ROBL2 more highly expressed in testis. Antibodies to ROBL2 label sperm flagella, consistent with interaction with axonemal dynein. Both ROBL1 and ROBL2 appear to interact with cytoplasmic dynein, indicating that these proteins are subunits of multiple dynein isoforms.47 Screening the human genome sequence with Chlamydomonas LC7a, ROBL1, and ROBL2 identified four regions of homology, on chromosomes 20, 16, 12, and 18 (table 10). The locus at chromosome 20q11 encodes ROBL1 and has been curated as protein DNCL2A (NP_054902). The locus at 16q23 encodes ROBL2 and has been curated as protein DNCL2B (NP_570967). The loci on chromosomes 12 and 18 are likely pseudogenes as the regions of homology contain stop codons, and no ESTs corresponding to these genes are in the EST database. Phylogenetic analysis (not shown) did not clearly distinguish the closest interspecies homologues between Chlamydomonas LC7A and LC7B and human DNCL2A and DNCL2B; in any case, both DNCL2A and DNCL2B are candidates for being subunits of both human outer arm and inner arm dynein.
Dynein light chain 7 loci
Docking complex genes
DC1
Chlamydomonas DC1 is a coiled-coil protein.48 Defects in the Chlamydomonas gene encoding DC1 (ODA3) block assembly of the outer dynein arm and the outer dynein arm docking complex, leading to reduced beat frequency and slow swimming.
Searching the databanks with Chlamydomonas DC1 (AAC49732) identified only low significance matches to coiled-coil proteins such as cingulin and myosin. Thus, it does not appear that there are any potential human orthologues of DC1 in databanks at this time.
DC2
Like DC1, DC2 is a coiled-coil protein49 and mutation of the Chlamydomonas gene (ODA1) encoding this protein blocks assembly of the outer dynein arm and outer dynein arm docking complex, leading to slow swimming.
Takada et al49 identified two human proteins (BAB71448 and BAB71508) as potential orthologues of Chlamydomonas DC2 and named these HsDC2-1 and HsDC2-2. These two polypeptides are likely to be the N- and C-terminal ends of the same protein; only BAB71508 has been assigned an interim gene symbol (FLJ32926) and RefSeq number (NP_653178). Searching the databanks with Chlamydomonas DC2 (AAK72125) or the peptide that results from combining HsDC2-1 and HsDC2-2 identified a locus on chromosome 19 that encodes HsDC2-1 and HsDC2-2 and a locus on chromosome 12 that encodes a hypothetical protein called FLJ35843 (NP_689804) (table 11). FLJ23926 (HsDC2-1+HsDC2-2) is 25% identical to Chlamydomonas DC2 over 488 residues (BLAST E value of 5e-34). FLJ35843 is 26% identical to Chlamydomonas DC2 over 403 residues (BLAST E value of 3e-31). FLJ32926 and FLJ35843 are only slightly similar to one another (24% identity over 220 amino acids, BLAST E value of 7e-15).
Dynein DC2 loci
Searching the non-human databases with these sequences identified a number of matches in lower eukaryotes as previously reported49 but also identified a significant match to an unpublished Ciona protein called axonemal p66 (BAB88833) as well as uncharacterised proteins from Xenopus and other vertebrates. Interestingly, Chlamydomonas DC2 is distantly related to the ChlamydomonasODA5 gene product (23% identical over 413 residues, BLAST E value of 9e-22). ODA5 will be discussed below.
DC3
Chlamydomonas DC3 (AAP49435) is an EF-hand protein related to calmodulin and centrin.50 Searching the human genomic sequence with Chlamydomonas DC3 identified two regions of homology (E<0.001) on chromosome 13, and one region on chromosome 18. The two loci on chromosome 13 are most similar to calmodulins but are likely pseudogenes as they contain stop codons within the predicted regions of homology and are not represented by EST clones. The locus on chromosome 18 encodes centrin-1 (NP_004057). Thus, we cannot identify a definitive orthologue of Chlamydomonas DC3 in the human genome at this time.
Other genes
All of the proteins known to purify with the Chlamydomonas outer dynein arm have been cloned and sequenced and are discussed above. However, genetic analysis indicates that additional genes are required for assembly of the outer dynein arm. One of these, ODA5, has been cloned and characterised; its product is associated with an adenylate kinase (AK) and the product of another gene, ODA10, which has not been cloned.
ODA5
ODA5 (AAS10183) is a coiled-coil protein that associates with the flagellar axoneme independently of the outer dynein arm and the outer dynein arm docking complex.51 Mutations in ODA5 block assembly of the outer arm but not the docking complex; as a result, the mutants swim at approximately one third normal speed.15,51 The only significant match in the human genome is to the FLJ35843 protein that was identified above as a potential orthologue of Chlamydomonas DC2. The match between ODA5 and FLJ35843 (21% identical over 321 residues, BLAST E value of 4e-12) is less significant than the match between DC2 and FLJ35843 (26% identical over 403 residues, BLAST E value of 3e-31), indicating that FLJ35843 is more likely to be orthologous to DC2 than to ODA5.
ODA5 associated AK
Analysis of oda5 mutant cells identified an AK (AAS10182) that was reduced in the flagella of oda5 and oda10 mutant cells.51 AKs increase the efficiency of ATP utilisation by converting two molecules of ADP to one AMP and one ATP; the resulting ATP can then be used by an ATPase like the outer dynein arm. Because no mutations in the gene encoding this AK have been reported, it is not known if this protein is required for outer arm assembly; however, because it is likely to be part of an axonemal complex (the ODA5/ODA10/AK complex) necessary for outer arm assembly, it is included here.
The human genome encodes multiple isoforms of AK. The Chlamydomonas ODA5 associated AK is composed of three AK motifs. No human proteins have this structure, but human AK5-1 and AK5-2 have two AK motifs.51 Searching the human NR protein and nucleotide databases with the Chlamydomonas ODA5 associated AK identified AK1 (NP_000467) encoded on chromosome 9q34.1 and the two isoforms of AK5 (NP_777283, NP_036225) encoded on chromosome 1p31 as the best matches51 (table 12).
ODA5 associated AK loci
DISCUSSION
Human orthologues of Chlamydomonas outer dynein arm genes
A remarkable finding of the present study is that 12 out of 14 subunits of the Chlamydomonas outer dynein arm have readily identifiable homologues in humans (fig 3). This provides further evidence that the proteins as well as the structure of the “9+2” axoneme of cilia and flagella have been highly conserved throughout evolution, and confirms the value of Chlamydomonas as an excellent model system for elucidating the molecular basis for human diseases that affect cilia and flagella.
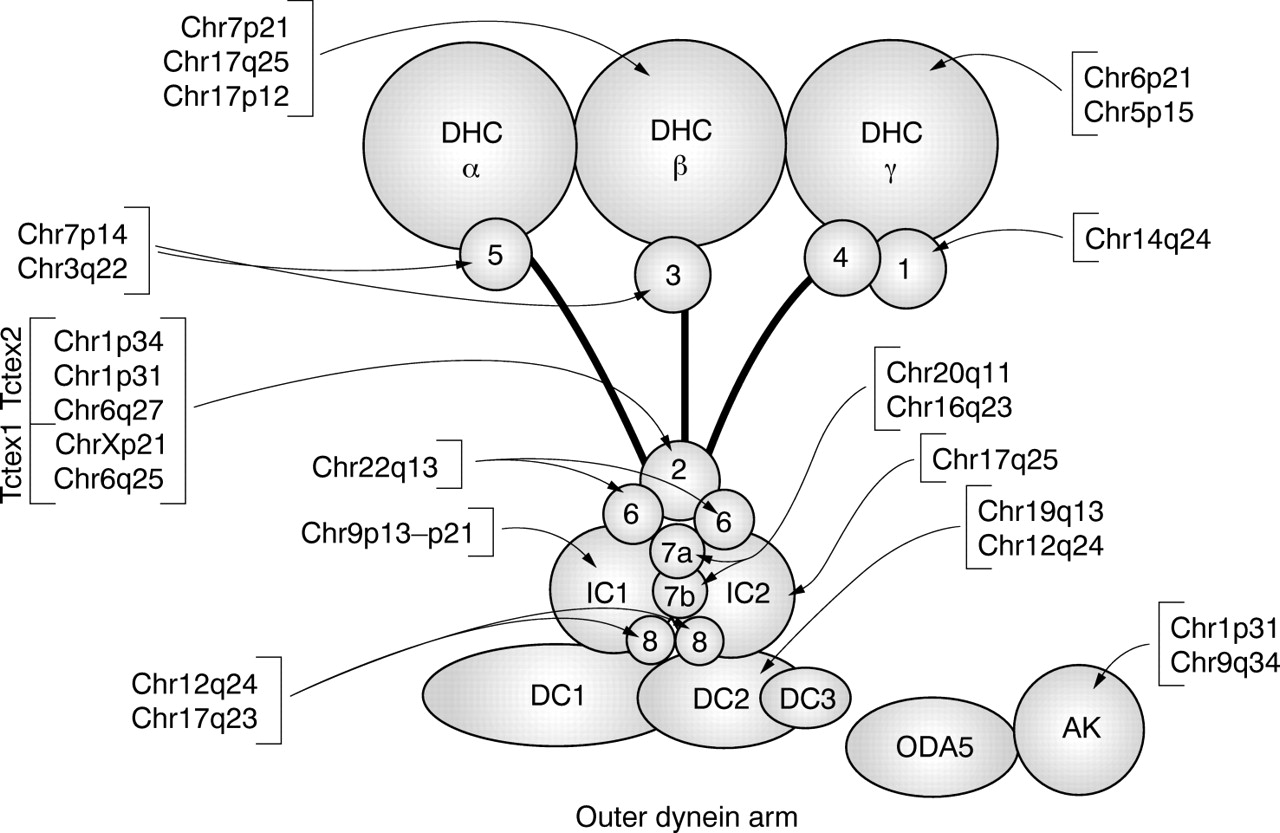
Human loci corresponding to the subunits of the Chlamydomonas outer dynein arm. Each of the loci identified in the human genome is linked to the corresponding subunit. Only genes that are likely to be expressed are included.
Another important finding is that several Chlamydomonas outer dynein arm proteins have multiple expressed homologues in humans (figs 3 and 4). This was particularly striking for the HCs: Chlamydomonas DHCβ has three close homologues in humans, and Chlamydomonas DHCγ has two close homologues. Inasmuch as the HCs are not known to be shared by different dyneins, it is likely that there exist multiple isoforms of outer arm dynein in humans. If some isoforms are expressed specifically in the testis and others in somatic tissues such as the lung, this could explain cases of human patients in which defects in respiratory tract cilia but not sperm flagella, or vice versa, have been reported.1 In addition, Chlamydomonas LC2, LC7a/b, and LC8 each have two or more close homologues in humans. In Chlamydomonas, these LCs or their close relatives are subunits of more than one type of dynein: for example, the LC2 relative Tctex1 is a component of inner arm dynein and cytoplasmic dynein,31,52 LC7a and LC7b are subunits of both inner arm and outer arm dynein,46,47 and LC8 is a subunit of outer arm dynein, the inner arm I1 dynein, and cytoplasmic dynein.31,42,53 Thus, it will be of interest to determine if the multiple LC homologues in humans represent evolutionary divergence and specialisation for use in different types of dynein, or if they function in different isoforms of outer arm dynein.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
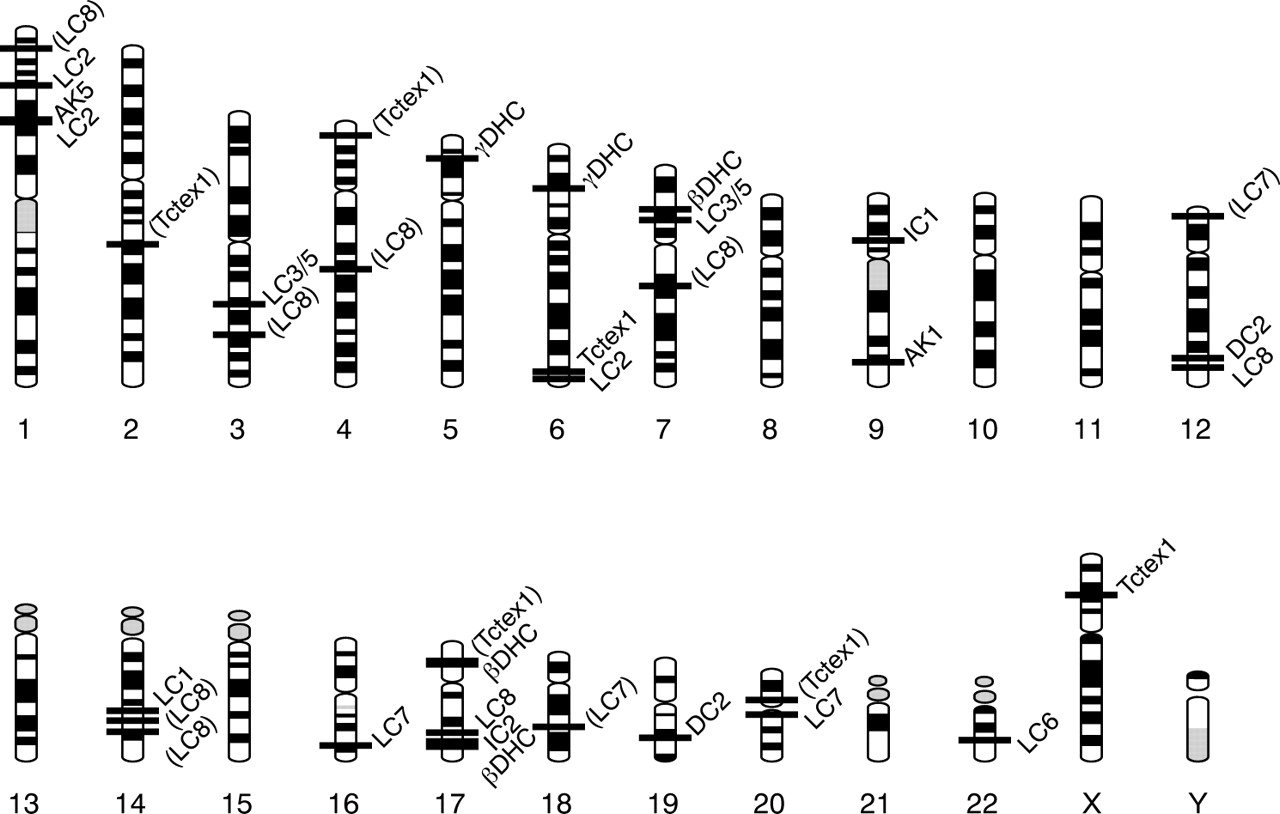
Chromosomal ideograms showing the approximate positions of all loci encoding potential outer dynein arm subunits. Parentheses indicate that the genes at these positions are not expressed and many are likely to be pseudogenes.
Outer dynein arm genes and PCD loci
Including the multiple human homologues of several Chlamydomonas outer dynein arm subunits, as well as two possible homologues each of DC2, the outer arm associated AK, and Tctex1, a total of 24 human genes that are potentially involved in outer arm assembly have been identified and mapped on the human genome (fig 4). These genes are good candidates to be screened for nucleotide changes in PCD patients with defects in the outer dynein arms. Three of these genes have already been shown to be defective in PCD patients (table 13). Pennarun et al3 sequenced exons from the human homologue (NCBI GeneID 27019 at 9p13-21) of the Chlamydomonas IC1 gene and identified two loss-of-function mutations in a PCD patient lacking outer dynein arms; further studies have led to the identification of additional mutations in this gene in affected individuals.54,55 Omran et al,56 studying a large family with PCD and absence of outer arms, mapped the PCD gene to a region (5p15-p14) containing a gene encoding one of the homologues of Chlamydomonas DHCγ. Olbrich et al4 then sequenced all exons of the candidate gene (NCBI GeneID 1767) in several affected individuals and found several mutations, most of which resulted in premature termination of the HC motor domain. A genome-wide scan of 70 affected individuals from 31 families identified several “suggestive” PCD loci57 (table 14), including one at chromosome 7p15.3-21 in families with dynein arm deficiencies; this region contains a gene encoding one of the outer dynein arm β HC isoforms (NCBI GeneID 8701), which Bartoloni et al5 subsequently showed to contain a nonsense mutation on both chromosomes in a patient with probable Kartagener syndrome but no obvious ultrastructural defects in the axoneme. Similar screens have been carried out on the genes encoding IC2 (GeneID 64446 on 17q25),25,26 another isoform of the outer dynein arm β HC (GeneID 1765 on 17p13),58 an isoform of TCTEX1 (GeneID 6991 on 6q27),59 and an isoform of LC8 (GeneID 140735 on 17q23.2),26 but did not identify mutations in these genes (table 13); however, failure to find mutations does not exclude the genes from consideration as candidate PCD genes in other PCD families. The work presented here identifies a number of additional candidate genes that should be considered in screening for potential PCD genes. Focusing on homologues that show outer dynein arm assembly phenotypes in Chlamydomonas (table 1) is likely to increase the success rate.
Outer arm dynein genes screened in human PCD populations
PCD loci
In addition to the three PCD genes noted above, numerous other potential PCD loci have been identified (table 14). The use of large families and isolated populations has identified PCD loci at 15q13-q15, 16p12,60 19q13-qter61 and X.62 The loci on chromosomes 16 and 19 are associated with outer dynein arm defects, whereas the patients with mutations on chromosomes 15 and X have defects in inner dynein arms. Our work has not identified any outer dynein arm genes at 16p12 that would be candidates for a PCD gene. An outer dynein arm docking complex protein is encoded at 19q13 but apparently lies just outside of the critical region (H Mitchison, personal communication). The locus on the X chromosome was found in a large Polish family that had retinitis pigmentosa associated with PCD.62 Interestingly, there is an isoform of TCTEX1 encoded on the X chromosome. Since Chlamydomonas Tctex1 is a subunit of inner arm dynein31 and a TCTEX1 subunit has been implicated in rhodopsin transport in retina,63 this gene should be considered a potential candidate in these patients.
The genome wide scan of Blouin et al57 identified “suggestive” PCD loci on 4q, 5p, 8q, 16p, and 19q with “potential, interesting” loci at 3p, 7p, 10p, 11q, 13q, 15q, and 17q; about half of the families in this study had defects in outer and/or inner dynein arms, whereas the other half had other abnormalities in ciliary structure or no apparent abnormalities. As noted above, the β HC, IC2, and LC8 genes on chromosome 17q subsequently were examined in some of these families and other PCD patients, and no mutations were found. The locus at 3p was identified by linkage analysis using all PCD families; therefore, the inner dynein arm HC genes DNAH1 and DNAH12 at 3p21 and 3p21.1 should be considered candidates for this potential PCD locus. Blouin et al57 identified 16pter as a suggestive PCD locus based on analysis of families with outer and/or inner dynein arm deficiencies; although no outer dynein arm genes map to chromosome 16, the inner arm HC gene DNAH3 at 16p12 may be a candidate for this potential PCD locus if the defect involves loss of inner dynein arms.
There are potential PCD loci that do not correlate with chromosomal regions containing presumed outer dynein arm genes. However, there are additional, uncharacterised genes necessary for outer arm assembly in Chlamydomonas51; cloning and sequencing of these genes probably will provide additional candidate PCD genes in humans, some of which may map to the PCD loci that are not currently correlated with outer dynein arm genes. In addition, some of the PCD families used in the linkage studies of Blouin et al57 had defects in axonemal substructures other than the outer dynein arm; an analysis of Chlamydomonas radial spoke proteins (Yang et al, in preparation) and a proteomic analysis of the Chlamydomonas flagellum7 are likely to provide candidate genes for some of these PCD loci.
In summary, we have used information from the model organism Chlamydomonas, in which the flagellar outer dynein arm has been extensively characterised, to identify a total of 24 genes that are predicted to encode subunits of this important motor in humans. Fifteen closely related genes that are likely to encode human inner dynein arm subunits are also identified. The resulting genes are excellent candidates for PCD disease genes.
Acknowledgments
We thank Dr Hannah Mitchison for her continued encouragement of this project.
REFERENCES
Supplementary materials
The appendix is available as a downloadable PDF (printer friendly file).
If you do not have Adobe Reader installed on your computer,
you can download this free-of-charge, please Click hereFiles in this Data Supplement:
- [view PDF] - Heavy Chains.
Footnotes
-
Published Online First 3 June 2005
-
This work was supported by grants from the NIH (GM-60992 to GJP and GM30626 to GBW), by a Worcester Foundation for Biomedical Research Foundation Scholar Award (to GJP), and by the Robert W. Booth Fund at the Greater Worcester Community Foundation (to GBW)
-
Competing interests: none declared